
要回答这个问题,我们需要回到前寒武纪时代。
7 亿年前,人和虫子们有着共同祖先。
这个共同祖先,长得类似于水母的浮浪幼虫[1]。
这个时期的动物,大家都只有一个口。
这个口,既可以作为吃东西的嘴,也可以作为排遗的肛门。
此时简单的消化道,就像袋子一般的简单腔体,因此也被称为腔肠。
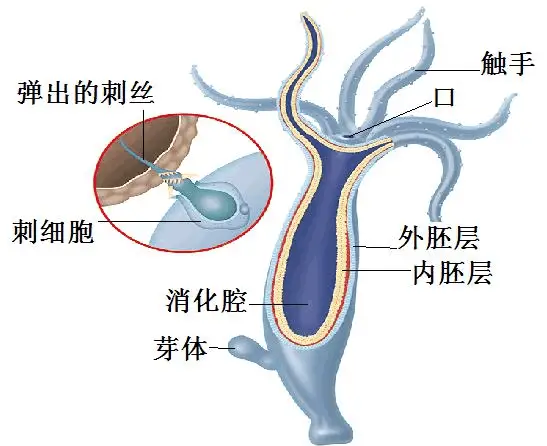
无论口的摄食效率,还是腔肠的消化吸收效率都太低了。
因此随着身体结构演化得越来越发达,因此有了专门开一个口的必要。
这个图中,长得像钵盂的,就是共同祖先了。
之所以开口这么大,其实主要还是过滤浮游生物、有机质,混口饭吃。
6.5 亿年前,另外一个开口的出现,真正意义上的肠道也随之出现。
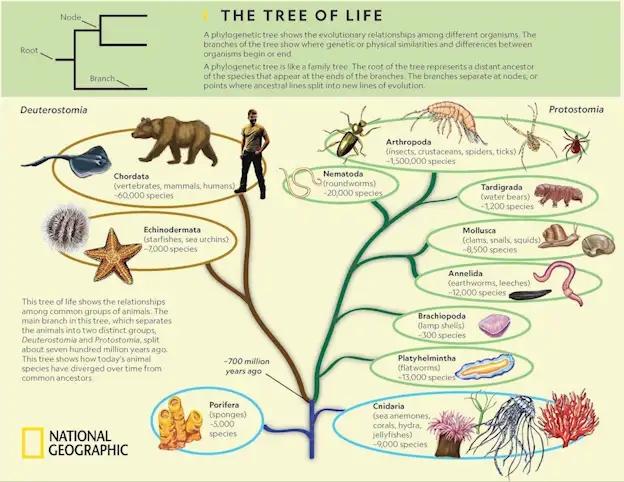
原来的开口,继续作为嘴巴,后来的开口作为肛门的,就是原口动物。它是扁形虫、蚯蚓、蜗牛、昆虫、蜘蛛等动物的共同祖先。
原来的开口,继续作为肛门,后来的开口作为嘴巴的,就是后口动物。它是海鞘、海星、文昌鱼,以及其它所有脊椎动物的共同祖先。
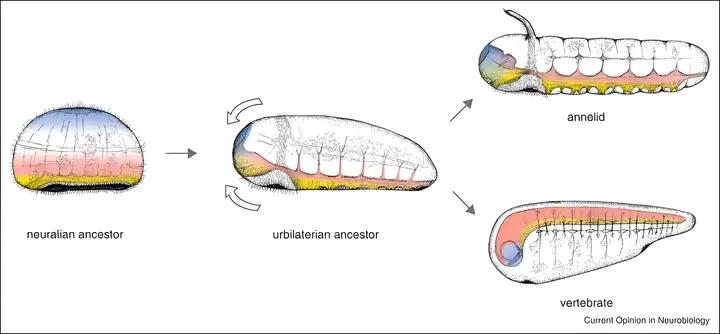
共祖的神经系统,是网状神经系统。就像水母一样,触摸一下,会有全身性的反射。
随着演化出原口动物和后口动物,神经系统也各自走上了集中化的道路[2][3]。
其中原口动物的肠道,向上穿过神经系统,具有发达的腹部神经索。
而后口动物的肠道则下行于中枢神经,后来演化出发达的脑脊神经系统。
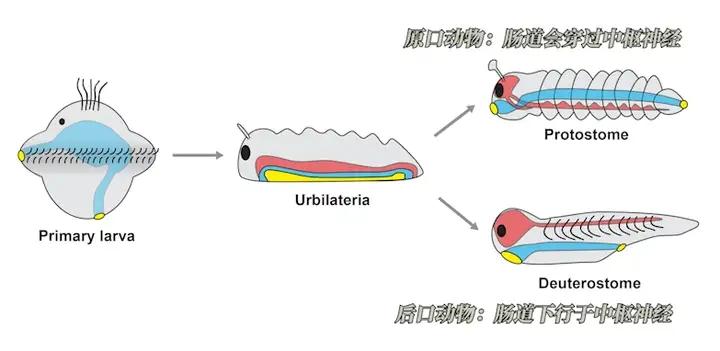
从此以后所有的动物都发展成了两侧对称,因此又称为两侧对称动物。
在这个过程,它们身体的内外两个胚层之间,也发展出来全新的胚层——中胚层。
身体多了一个胚层,也就意味着原来的肠道和外壁之间出现了全新的腔体——体腔。
中胚层的形成方式,有裂体腔法和肠体腔法。
绝大部分的原口动物都是裂体腔,绝大部分的后口动物都是肠体腔。
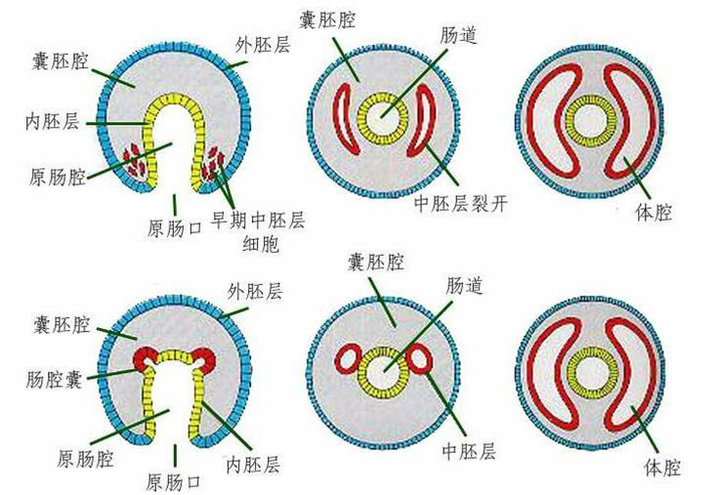
体腔的出现,也意味着早期动物身体结构出现了前所未有的变化,不仅有了独立的肠道,还发展出了各种各样的器官。
这些和哺乳动物 / 昆虫的血肉之躯 / 汁水之躯有什么关系呢?
因为原口动物和后口动物刚刚分化出来的时候,和早期祖先一样,都没有循环系统。
这个时期,由于个体小,再加上新陈代谢还比较弱,O₂和 CO₂的交换,营养物质的运输,代谢废物的排出,仅仅通过简单的扩散作用,就足够了。
然而 5.42 亿年前,进入寒武纪以后,原口动物和后口动物的身体结构变得越来越复杂,单纯通过简单的扩散作用,已经不再能提供 O₂和 CO₂的交换、营养的运输,以及代谢废物的排出。
动物身体在满足机体新陈代谢的过程中,自然而然也就演化出了循环系统。
节肢动物开放式循环系统的演化。
早期演化出来的泛节肢动物,就已经发展出了外骨骼。
外骨骼与体腔,可以直接形成一个稳定的“体液池”。这样的情况下,只需要发展出一个类似于心脏的结构,驱动这个液体池循环,就可以完成体液循环。
这便是节肢动物的开放式血液循环——血淋巴循环。
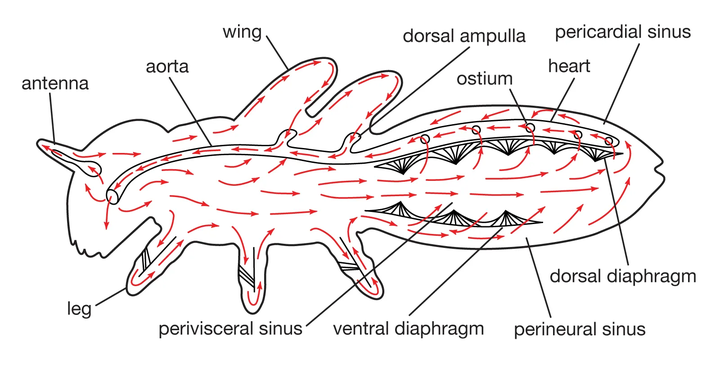
通常来说,对于绝大多数节肢动物来说,氧气的交换,直接通过鳃扩散就足够了,无需专门携带氧的蛋白。
但虾、蟹等多数甲壳类,以及鲎、蛛、蝎等蛛形纲成员,这些节肢动物要么体型大,要么活跃,要么生存于低氧 / 低温环境,对氧气的运输能力有着更高的需求。
因此发展出了,与脊椎动物类似的蛋白携氧机制。
它们身体内存在血蓝蛋白(hemocyanin),来协助氧气的运输。这种蛋白脱氧状态为无色,结合氧后,因铜离子的氧化而呈蓝色。
相比起血红蛋白,它们的开放式循环,更适合血蓝蛋白。因为这种蛋白是直接溶解在血淋巴中的,也正因为如此,它们没有演化出专门运输氧的红细胞。
但它们身体内还是具备多种血细胞的,可负责免疫、修复、吞噬、凝聚等与脊索动物相似的多种功能。
其实,称呼这类节肢动物的血淋巴为血液,也并不算不妥。

后来,一支登陆的节肢动物演化成了昆虫。
它们高度适应陆生环境,发展出了全新的呼吸系统——气门系统。
气门系统,直接把氧气扩散到身体的各个角落。
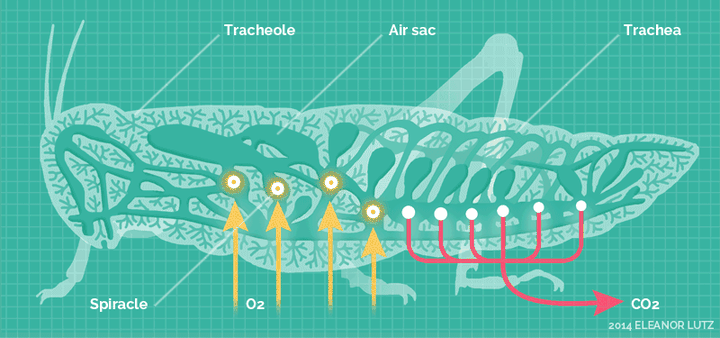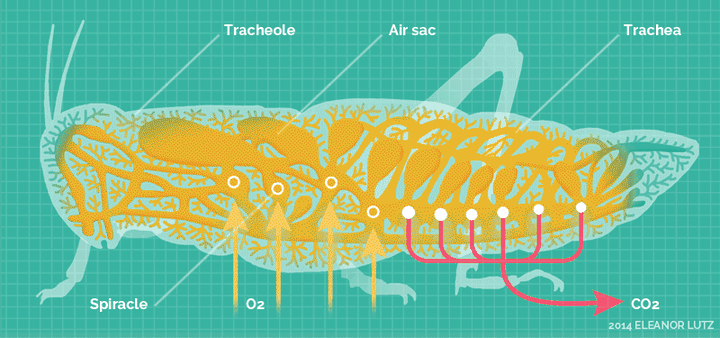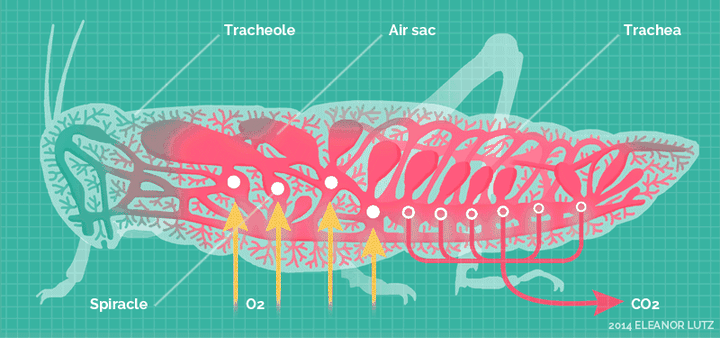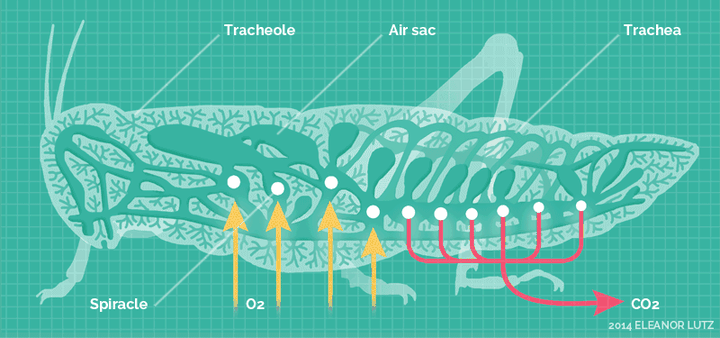
这套呼吸系统,对于昆虫的体型来说,非常的高效,使得它们不再依靠祖先的血蓝蛋白。除了少数昆虫,绝大多输都不再具备血蓝蛋白表达。
因此对于昆虫来说,它们的血淋巴除了不具备氧气携带能力外,其它功能和脊索动物这一支的都非常相似。
相比起一些海洋节肢动物和脊索动物用葡萄糖作为血糖,昆虫并不如此。
它们使用海藻糖(trehalose)作为血糖。
海藻糖比起葡萄糖更加的稳定,对于它们小体型身体去适应各种严苛、多变的环境,具有非常大的优势[4]。
世界上能脱水“复活”的动物,多以海藻糖作为血糖。
轮虫纲——蛭形轮虫,从数万年永冻土中分离复苏[5] [6],能盗取基因[7],约 8~9%外源基因。
线虫动物门——秀丽隐杆线虫,耐久型幼虫阶段可脱水 98%[8],可在极端干燥环境存活 4 个月。
缓步动物门——水熊虫[9],可脱水 99%,抗极端高低温、辐射,存活数年,有盗取基因能力。
鳃足纲——丰年虾[10],俗称卤虫,耐干旱高盐,幼虫脱水 98%,最高可耐 100%盐浓度。
昆虫——范氏多足摇蚊[11] [12],又称昏睡摇蚊,脱水 97%,长时间耐高低温,耐纯酒精。
脊索动物封闭循环系统的演化。
后口动物这一支,早期并没有走上叠甲这一条路。
大多是软体的,没有明显外骨骼。
它们早期发展出来的循环系统,只能在身体内部开辟出管道,来把氧气、营养输送到全身。
但一开始这些管道内的物质,并不能很好的与身体组织隔绝,因此属于半开放循环系统。
这样的循环系统,无论氧气还是营养物质、代谢废物的交换效率,都会比较低。
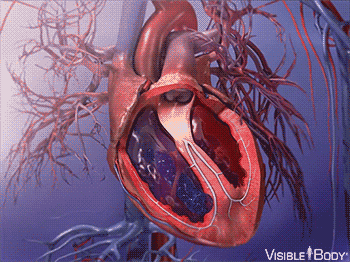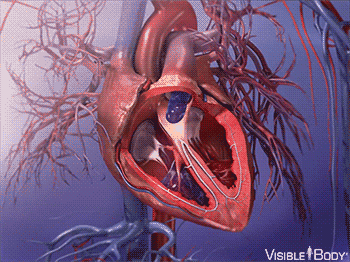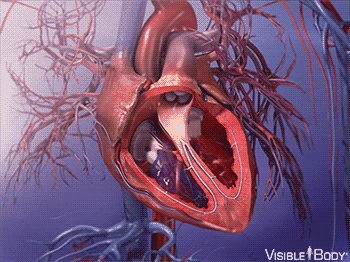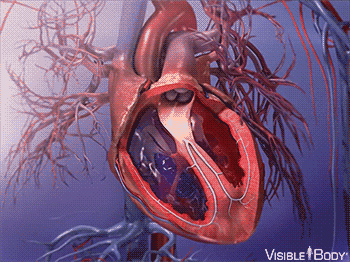
因此,大约在 5.4 亿年前,最早脊索动物诞生的前后,它们发展出了一种特殊的内皮细胞——血管内皮细胞。
这种细胞,最终发展成了血管,使得脊索动物都有了专门的封闭管道,来输送血液。
同时,腹部大动脉发展出搏动能力,血液循环了起来。
在后来漫长的演化过程中,这个拥有搏动功能的腹部大动脉,最终发展成了有着复杂结构的心脏。
脊索动物之所以最终选择了血红蛋白携带氧气,演化压力主要来源于巨大的需氧量。
同时发展出专门运输氧气的红细胞,再把血红蛋白封装在里面,既可以高浓度运载氧气,又能保护机体免受游离血红蛋白的毒性损伤。
在血糖的选择上,脊索动物之所以选择了葡萄糖,同样和封闭系统的高能量需求有关。
昆虫的海藻糖,虽然在稳定性上非常具有优势。但并不能被细胞直接利用,需要先水解成葡萄糖,才能进入能量代谢途径。
这其实有着更高的代谢成本。
脊索动物直接利用葡萄糖,自然是最直接、也是效率最高的供能方式。
尤其是在后来的演化过程中,随着身体、器官结构越来越复杂化,体型越来越大型化,巨型化。封闭的血液循环、红细胞、葡萄糖血糖等基本配置,都成了这些复杂演化的基石。
总的来说,哺乳动物是血肉之躯,昆虫是汁水之躯,早在 6.5 亿年前就埋下了伏笔。早期演化上的小小歧,一步步造成了今天的巨大差异。