


不是不想,是臣妾做不到啊!
20 亿年前的老祖宗,哭晕在厕所。
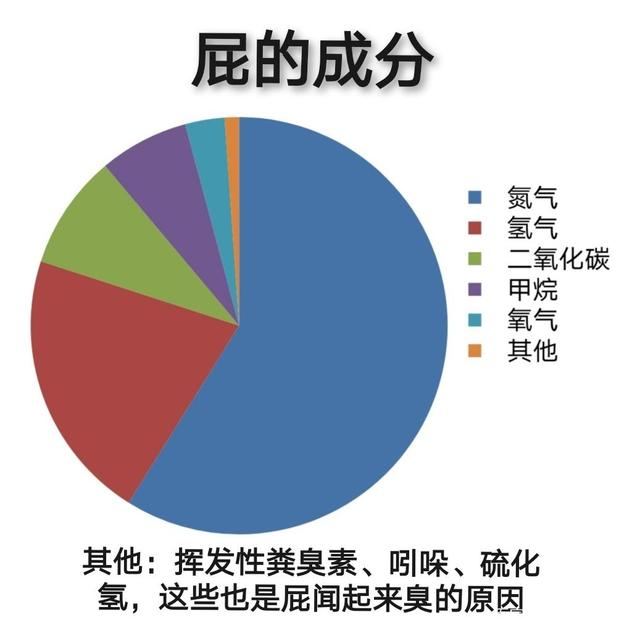
你以为只有哺物动物不会利用甲烷,实际上所有动物都不会。
不仅仅是所有动物不会,所有真菌、植物,乃至于整个真核生物都不会。
也就是说,从 LUCA 到我们这一支,除了细菌、古菌,至少长达 20 亿年都没有甲烷代谢的能力。
不说甲烷了,知道金针菇吧?
你吃了金针菇,会原封不动拉出来,是金针菇不够营养吗?

人类无法消化金针菇,是因为金针菇细胞壁富含几丁质和纤维素。
几丁质是一种含氮多糖。
你看看这化学式,不够营养吗?
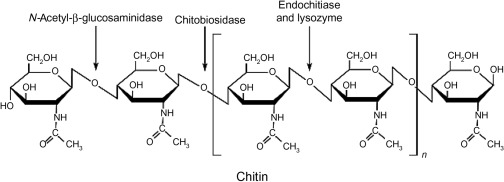
几丁质广泛存在于各种无脊椎动物的体表。

如果人类能消化几丁质,虾蟹贝都可以是更高能量食物。
哺乳动物身体内主要就壳三糖酶(Chit1)和酸性哺乳动物几丁质酶(AMCase)具有一定消化几丁质的能力,但消化能力非常的有限,都不足以高效降解大分子几丁质[1]。
金针菇所含的纤维素,有一半是不可溶的[2]。

纤维素是由葡萄糖组成的大分子多糖。
你看看这化学式,不够营养吗?
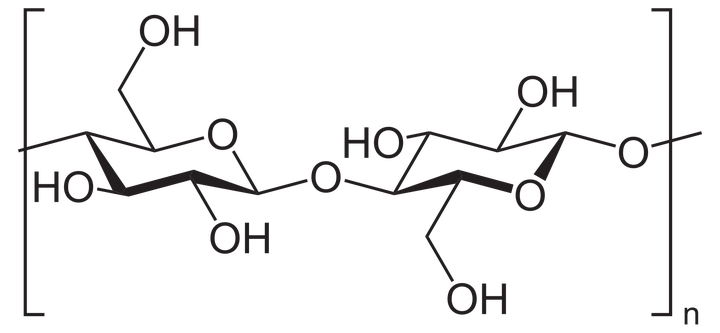
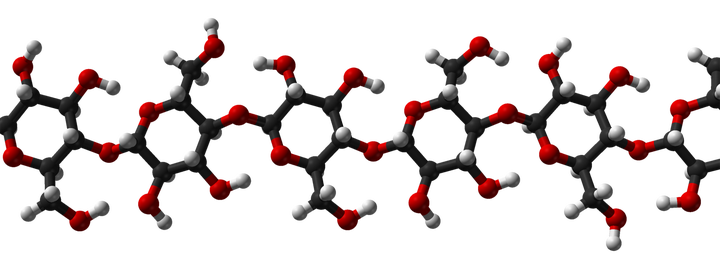
所有哺乳动物身体内,都缺乏纤维素酶。
仅仅金针菇内的大分子聚合物人类无法处理吗?
并不是。
甘露醇(Mannitol)[3]够简单了吧。
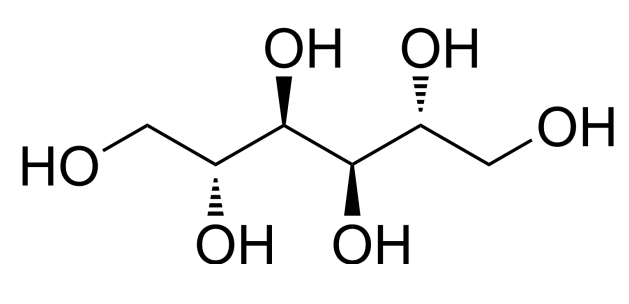
化学式
,仅仅比葡萄糖(
)多两个氢。
但人类就偏偏没有相关的呼吸链,来代谢它。
海藻糖(trehalose)[4]也够简单了吧?
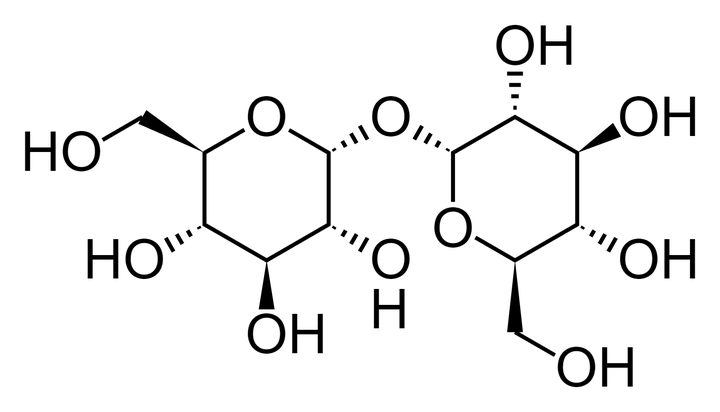
化学式
,和蔗糖的化学式一模一样(分子结构不同)。
海藻糖在无脊椎动物体内普遍存在[5],以血淋巴糖的形式存在,也即相当于我们的血糖。
哺乳动物中,除了食虫蝙蝠等专性食虫的类群海藻糖酶基因保留完整外,绝大多数都不完整,消化海藻糖的能力有限。
显然在自然选择面前,生物演化不能想要又要。
动物祖先选择了最后都容易分解成小分子的三大营养物质,那么在一次次优化迭代的过程中,各种有机聚合物的消化能力,自然而然就优化掉了。
除了真菌与部分原生生物,具备完整的内源性纤维素酶体系,能直接将纤维素水解为单糖。所有动物自身都缺乏高效纤维素酶。
自身无法利用,显然就只能利用外物了。
从细菌、古菌处“拿来主义”,至少可以追溯到 20 亿年前的内共生起源。
部分无脊椎动物通过水平基因转移,获得内源纤维素酶,而共生微生物发酵则是大多数能消化纤维素的动物所选择的方式。
牛羊和马驴能消化纤维素,正是因为它们都是借助肠道菌群来消化。
牛羊为前肠消化动物,马驴为后肠消化动物。
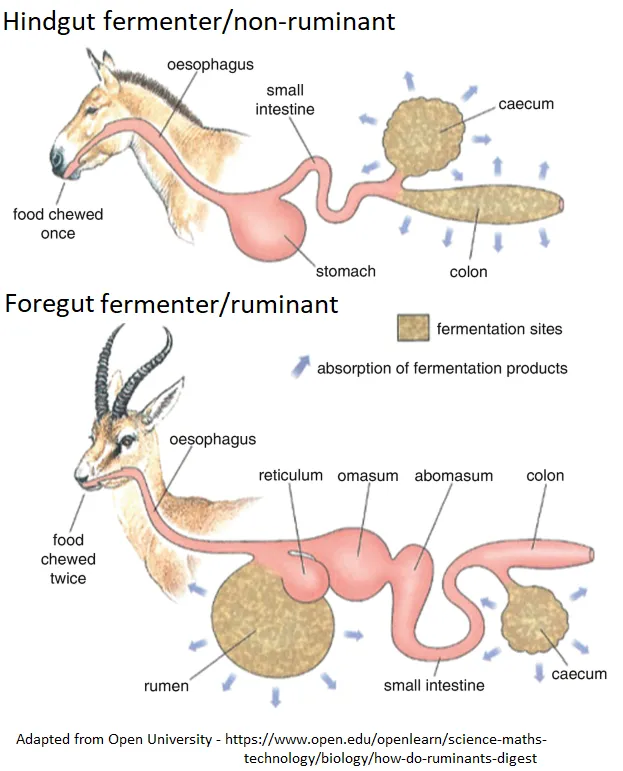
前肠消化动物主要是反刍动物[6]。
反刍动物的瘤胃内存在大量微生物,包括纤维分解菌和产甲烷菌。
这些微生物在分解植物纤维素等碳水化合物时,会产生氢气和二氧化碳。
产甲烷菌利用这些气体,通过生化反应生成甲烷,并通过打嗝或排气的方式将其排出体外。
显然你已经发现了,反刍动物体内的菌群甚至能利用氢气,但依旧无法利用甲烷。
目前只有一些细菌和古菌可以利用甲烷,统称为甲烷氧化菌(Methanotroph)[7][8]。
真核生物起源于 20~27 亿年前。
细菌起源于 25~35 亿年前。
古菌起源于 31~41 亿年前。
如果说,从 LUCA 到我们这一支,直接消化纤维素的能力已经丢失了 15 亿年。
那么代谢甲烷的能力,就至少丢失了 20 亿年,甚至可能长达 30 亿年。
甲烷是一种化学性质极为惰性,且能量密集的气体,要对其有效代谢,需要专门的甲烷单加氧酶(methane monooxygenase)[9]等关键酶系。
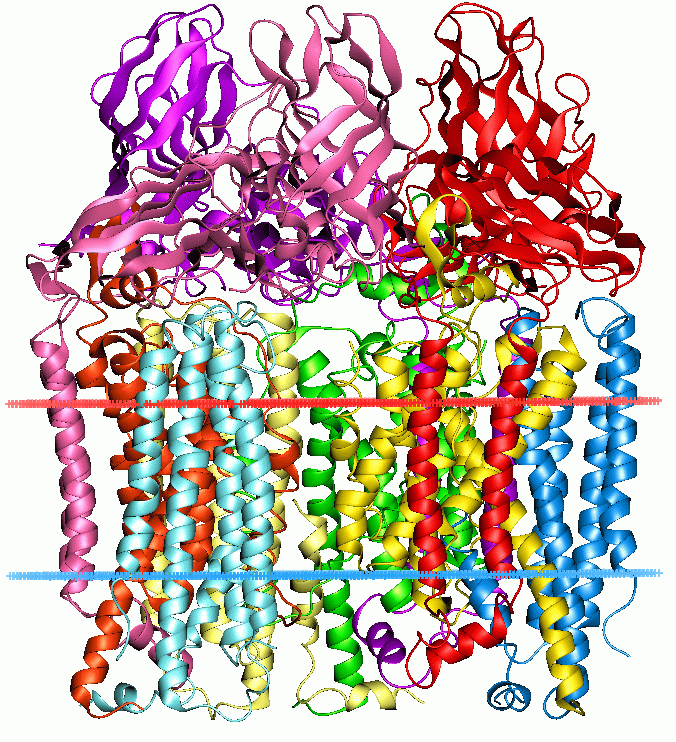
这些酶仅发现于甲烷营养菌、甲烷氧化古菌等特殊微生物体内。
它们依赖复杂的金属辅基(铜中心或二铁中心)和专门的电子传递链,才能在相对温和条件下活化 C–H 键。
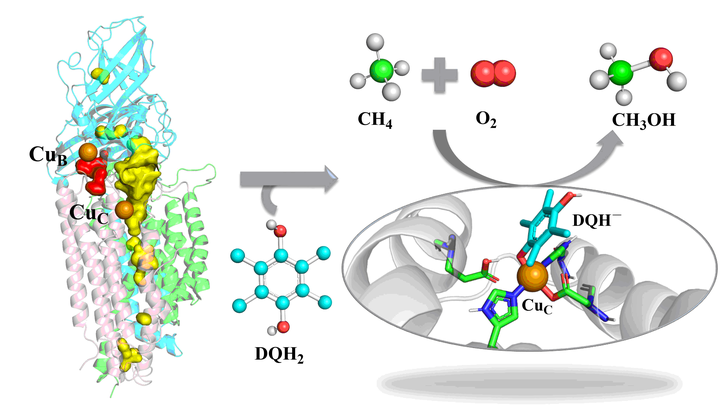
毕竟碳氢键的键能比较高,达到 435 kJ/mol。
而且代谢还不能一步到位,甲烷先转化为甲醇后,还需依次通过甲醇脱氢酶、甲醛脱氢酶等等进一步代谢过程。
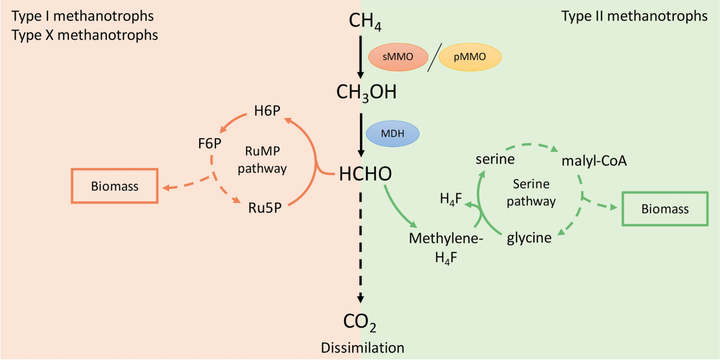
动物要有这样的一系列代谢能力,不仅需要相应的一系列基因合成甲烷单加氧酶,还需要相应的金属酶合成、调控网络。
这个过程,细胞需提供大量蛋白质的合成与更新,涉及大量 ATP 的消耗与补充。
更何况,造成整个代谢过程,还需要演化出,类似线粒体或叶绿体这样的专用细胞器,来隔离高氧化活性的中间产物,不然会对 DNA 和蛋白质造成严重的氧化伤。
说得更加直白一点,直接一个甲醇、甲醛,你就受不了,直接交命投降。
根据不同人的耐受能力,甲醇致死水平波动很大,15.8~474g 都有,中位数 56.2g[10],多数人在 1g/kg[11]。甲醛 (福尔马林)的致死数据没有甲醇丰富,最常用数据为 30ml 的 37%浓度福尔马林达到致死量[12],也即致死剂量大约在 10g 左右。
显然,如果没有额外的保护机制,大量代谢甲烷,对动物是极其致命的。
而产甲烷菌则有专门的胞内囊泡来隔离这些中间产物,从而避免代谢毒性的威胁[13]。
那可不可以继续拿来主义呢?
当然是可以的。
反刍动物瘤胃内的确有甲烷营养菌,它们能吸收利用其代谢产物,转化为短链脂肪酸(SCFAs)[14]。
那为什么这种模式没有成为主流,利用所有的甲烷呢?反而绝大多数甲烷都被打嗝排放了呢?

本质上还是哪怕甲烷共生代谢,也比较鸡肋。
毕竟动物获得营养的途径多种多样,绝大多数碳水化合物、脂质、蛋白质都是肠道直接消化吸收利用,并无迫切需求用甲烷功能。
更何况,甲烷在动物活动中虽然普遍存在,但浓度低且不稳定,难以成为可靠能源,无法获得优势。
即便能稳定获得,随着利用多了,代谢毒性成本也完全受不了。
相比之下,甲烷营养菌生活在高甲烷环境,获得的能量可以显著高于各种形式的成本,从而在相应生境中具有生存优势。
总之,动物体内缺乏甲烷代谢能力,主要在于相关酶系专属性强、完整代谢途径复杂、代谢成本高昂、生理风险高、生境压力不足等多种原因共同促成的。