
得到了较为稳定的酶活性。
我主要是来补充一个“失去了什么”的——失去了心脏的快速再生能力。

心肌梗死的预后问题是个困扰医生和病人很久的问题,这个事就是失去了心脏快速再生能力的结果,而这个结果就与恒温有关,甚至可以说是恒温的代价。
很多人忽略了一个问题:恒定的体温不是类似你用空调一样直接一按按钮体温就乖乖保持正常体温范围内的,它是无时无刻不需要精确微调的,调控体温的若干种激素中包括了一种含碘的激素——甲状腺素。
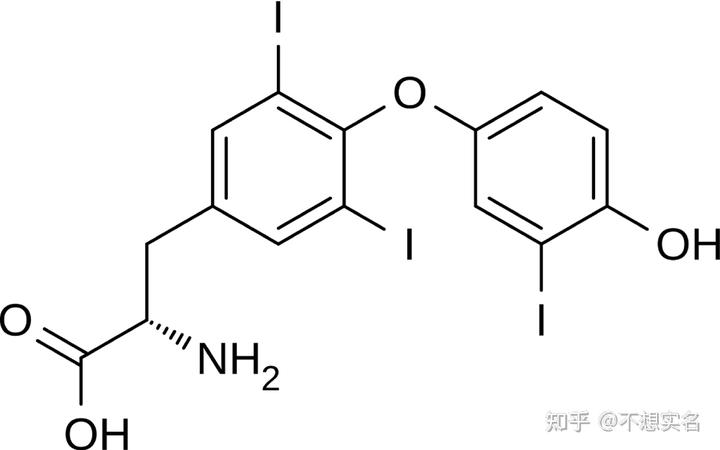
在体温调节中,寒冷信号会刺激下丘脑释放 TRH(促甲状腺激素释放激素),然后就是选生物的高中生都能说的步骤:TRH 在垂体发挥作用,使垂体释放 TSH(促甲状腺激素),TSH 作用于甲状腺,使其释放甲状腺激素,T3 甲状腺素与其受体结合,作为转录因子上调线粒体解偶联蛋白UCP1的表达(实际上并不完全是这样,可以去后面看 T3 甲状腺素受体是如何作为转录因子调控基因表达的),解偶联蛋白作为质子通道使被复合体 I、III、IV 传递出去的质子不经 F1F0-ATPase 就迅速回流至线粒体基质,从而使氧化磷酸化解偶联,氧化产生的能量变成热能,维持了体温——这就是甲减患者体温偏低的原因——而能够维持体温就意味着体内甲状腺素含量较高,能够在体温下降时及时使氧化磷酸化解偶联。
那么,体内甲状腺素含量较高的代价是什么呢?
成体哺乳动物的心肌细胞是多倍体(心肌细胞 90%以上为四倍体甚至更高),而幼年哺乳动物和冷血动物(如斑马鱼)的心肌细胞则多为二倍体,而无论是形成多倍体的过程还是结果都会造成细胞分裂受到抑制。
首先,形成多倍体的过程就是使 DNA 复制与细胞分裂解偶联的过程:一个细胞分裂期之前存在一个细胞分裂的间期,间期又分为 G1、S 和 G2 期,其中只有 S 期存在 DNA 的复制,而每一个时期到下一个时期(包括 G1 到 S、S 到 G2 和 G2 到分裂期,分裂期内还有若干检查点)的切换都需要通过细胞分裂检查点,满足分裂条件并通过检查点后才能进入下一阶段,这一过程中周期蛋白依赖性蛋白激酶(cyclin-dependent kinase,CDK)起到了很大的作用。
CDKs 为一组对真核生物的细胞分裂具有重要调控作用的丝氨酸 / 苏氨酸蛋白激酶,人体内共有 7 种 CDKs,它们通过与不同周期蛋白(Cyclin)组成有活性的酶,调节细胞周期的各个阶段转化,各种 CDKs 对应的 Cyclin 伴侣及其作用见下表:
| CDK | 与之互作的 Cyclin | 复合物作用 |
|---|---|---|
| CDK1 | Cyclin B | G2 期向 M 期转化 |
| CDK2 | Cyclin A | S 期向 G2 期转化 |
| CDK3 | Cyclin E | G1 期向 S 期转化 |
| CDK4 及 CDK6 | Cyclin C | G0 期向 G1 期转化、G1 期向 S 期转化 |
| CDK5 | Cyclin D | G1 期向 S 期转化,参与 Rb(视网膜母细胞瘤)基因产物的磷酸化 |
| CDK7 | Cyclin H | 与 MAT1 组成三元复合物(CDK 激活激酶复合物,CAK),激活其他 CDK,推动细胞周期进程 |
细胞分裂过程中存在一系列检查点,只有通过这些检查点,对应的 CDK 才会与 Cyclin 形成复合物,从而使细胞分裂进行下去。
那么有小伙伴就要问了:这和甲状腺素有什么关系呢?
当然有啦,甲状腺素可以通过多种机制抑制 CDK1/Cyclin B 复合物的活性并降低其含量,从而抑制细胞从 G2 期向 M 期的转化,使心肌细胞形成多倍体,随后该抑制作用扩展至更广泛的 CDK/Cyclin 复合物,造成心肌细胞不能进入细胞周期,并导致了心脏中不存在类似骨骼肌中卫星细胞的肌肉干细胞,使心脏成为了不能再生的器官。这一现象得到了各种实验证据的佐证:出生时小鼠血清中甲状腺激素含量较低,但在出生后短时间内迅速升高(可达原本的 50 倍),而正是在这一时段其心肌细胞大量多倍化,而敲除 TRα1 及使用甲状腺素合成抑制剂的小鼠心脏中多倍体细胞数量大幅度减少,并且保留了部分分裂能力,这说明其抑制作用主要由心肌细胞中的TRα1介导的。

TRs 为一组 2 型核受体(是的,这玩意在细胞质和细胞核中,而细胞外的甲状腺激素需要由 T3 转运体运进来才能起作用),其存在锌指结构,能够与 DNA 结合并作为转录因子。根据受体的位置,甲状腺激素进入细胞后分为两条通路:通路一由细胞质中的 TR 介导,通过上调 cGMP 的含量,激活 PKG,延长细胞的寿命;而通路二则是由细胞核中的 TR 介导,通过与结合有共激活因子的 RXR(Retinoid X receptor,类视黄醛 X 受体)结合为复合物发挥转录因子作用。
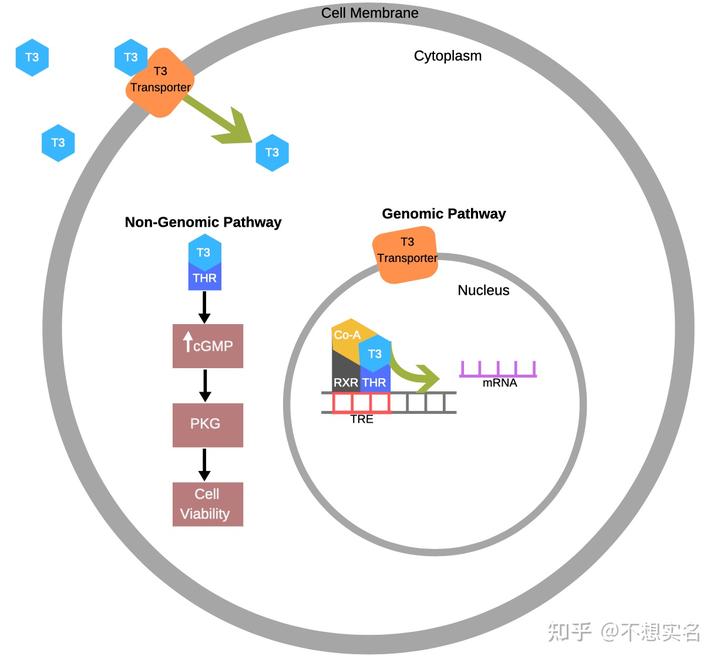
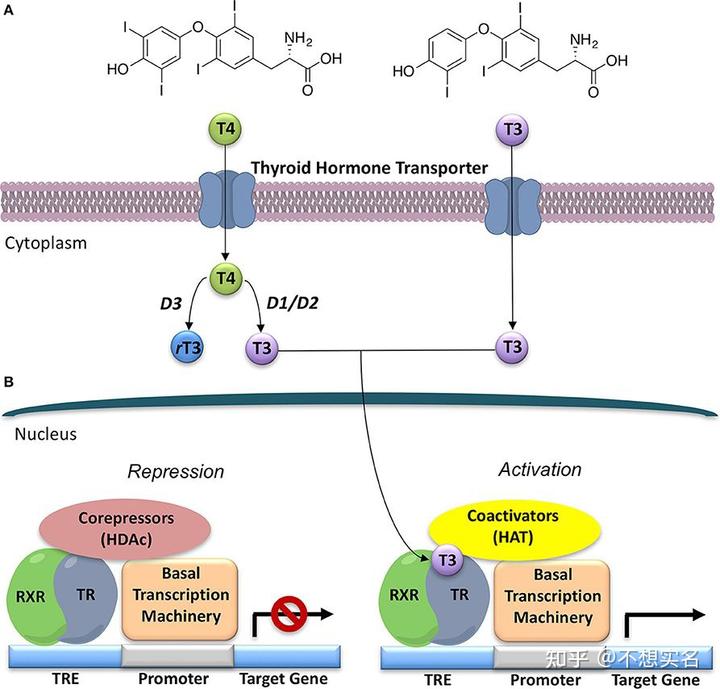
心肌中主要存在TRα1,该蛋白与 RXR 结合后形成的复合物的作用之一就是下调 CDK1 与 Cyclin B 的表达、上调 CDK3 与 Cyclin E 的表达,使心肌细胞多次进行 DNA 复制并停滞在细胞周期的 G2 期,这些都导致了心肌细胞的多倍化与分裂缓慢。
同时,多倍化的细胞可能被视为染色体突变或 DNA 损伤的产物,从而激活Chk1/Chk2,阻止 CDK1 与 Cyclin B 的结合,进而阻止 CDK1 活化,间接阻止了心肌细胞的分裂。
因此,我们心肌梗死后只能依靠成纤维细胞补上死亡的心肌细胞的位置,无法在短时间内再生新的心肌。
这大概就是此心唯一吧。