
有倒是有,但所谓重构也不过是从一座屎山,奔向另一座更大的屎山。不过,对于本文的主角,真核生物中最权威的屎山拥有者——甲藻(Dinophytes)来说,那是它生存的最终秘技。
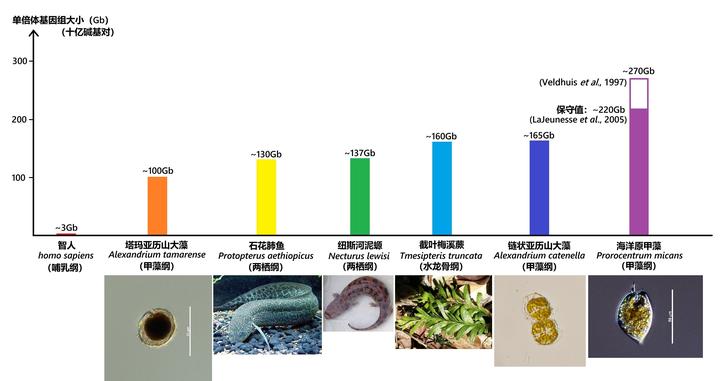
先来直观地感受一下甲藻的基因组有多恐怖。现在正在看这篇文章的你,作为智人(Homo sapiens)中的一员,单倍体基因组约有 30 亿碱基对,也就是3Gb;
动物中最知名的屎山代码,非洲肺鱼属的石花肺鱼(Protopterus aethiopicus),基因组有 1300 亿碱基对,即130Gb;
已知单倍体基因组最大的动物,泥螈属的纽斯河铌螈(Necturus lewisi),基因组高达137Gb;
已知基因组最大的植物,梅溪蕨属的截叶梅溪蕨(Tmesipteris truncata)略胜于前者,基因组大小来到了160Gb;
而甲藻家族中的最强者——海洋原甲藻(Prorocentrum micans),作为体长仅有 50 微米的单细胞生物,其基因组大小纵使按照比较保守的测定结果,都有220Gb,即 2200 亿碱基对。如此离谱的数值使它当仁不让地成为了已知单倍体基因组最大的生物,或许都没有之一。

看到这儿,相信你也会有疑惑:这 DNA 毕竟是实打实的,具有物理实体的物质,那甲藻到底是如何在一个娇小的身躯中塞下这么多基因的呢?
简单来说,是因为甲藻拥有一个在真核生物中最为特殊的细胞核,也就是所谓的甲藻核(dinokaryon)。曾几何时,生物学家对此困惑不已,因为甲藻核的结构实在是过于震撼,以至于有一种流行多年的假说,认为甲藻是真核生物中最原始的类群。该假说的支持者甚至还专门成立了介核生物(Mesokaryotic)这一演化支来容纳甲藻,意思就是介于原核生物与真核生物之间:
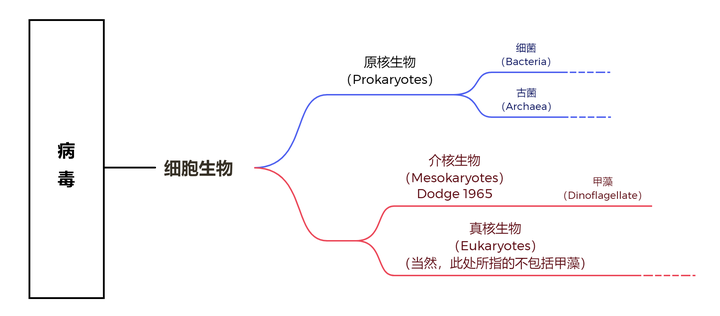
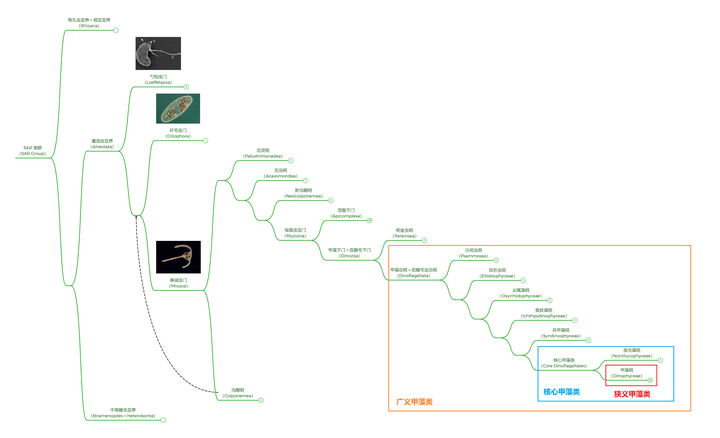
今天的我们会觉得这一假说简直荒谬到可笑,但在当时,这可是写进教科书的存在(骗你的,到现在也有一些教材是这么写的,那就是真的可笑了)。直到 21 世纪,分子生物学崛起之后,人们才逐渐对甲藻的演化位置达成共识。简单来说,甲藻是 SAR 超类群的成员,而与它关系最近的大类群是以疟原虫为代表的顶复类(Apicomplexa),再远一点就是以草履虫为代表的纤毛虫门(Ciliophora):
而甲藻核,可以说是真核生物历史上一次绝无仅有的细胞核重构带来的产物。这个故事开始于距今两亿多年前的三叠纪,当此之时,显生宙以来最惨烈的二叠纪末大灭绝刚刚过去,而曾经霸占海洋光合浮游生物生态位的红藻和绿藻都在这次事件中损失惨重。新一代的藻类眼看旧王退位,便纷纷趁势崛起;我们的主角甲藻自然也是其中的一员,只不过,命运很快就把它带上了一条未曾设想的道路。
在某一天,一个属于核质病毒门(Nucleocytoviricota)的巨病毒感染了甲藻的祖先;这本来也是一件再正常不过的事,但在一系列机缘巧合之下,这个病毒把一段编码甲藻病毒核蛋白(DVNP)的基因落在了甲藻祖先的细胞核中。可怜的甲藻估计也没想到,这段基因将开启它的登神长阶。
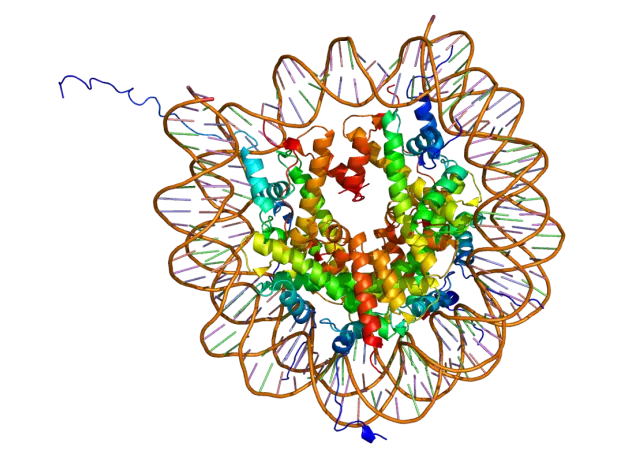
DVNP 有一个特别厉害的能力:它能将 DNA 压缩到极致,使其根本无法发挥作用。据推测,病毒演化出这种蛋白的本意是为了迫使宿主的抗病毒基因停摆,但在 DVNP 被甲藻夺舍了之后,情况便发生了微妙的变化。众所周知,真核生物本就内置了一套 DNA 压缩系统,由一类名叫组蛋白的碱性蛋白质完成,但它的压缩比和 DVNP 比起来根本不值一提。
这样一来,在细胞核有限的空间内就能放下更多基因了。什么,你说 DVNP 会导致 DNA难以被转录?没关系,毕竟在浩如烟海的基因中,真正工作繁忙的就没几个,而那些闲置的基因刚好可以用 DVNP 打包待机。于是,在此后的演化中,组蛋白在甲藻核内变得越来越少,直至被彻底抛弃。
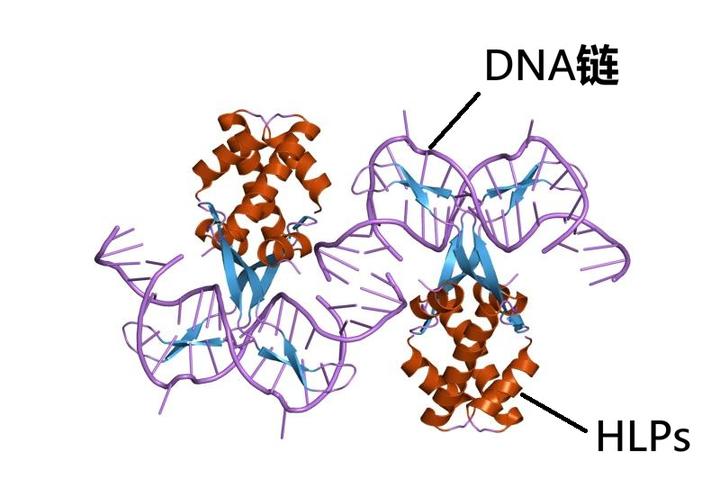
到了这一步,甲藻显然也没有了回头的可能,只能仗着高压缩比腾出的空间收集各路基因,以期找到能够为自己提供生存优势的片段。结果,还真就有一支甲藻从细菌那里捡到了宝藏。这段新的基因序列编码了一类被称为组蛋白样蛋白(HLPs)的蛋白质,它广泛分布于原核生物体内,但在真核生物这里却早已丢失。如今,这个被遗弃了 20 多亿年的老代码重新回到了甲藻手中,而它们很快就发现,这种蛋白质居然可以让那已经被 DVNP 压缩到惨不忍睹的基因重新转录出来,就像是一套与 DVNP 相配合的解压算法。这样一来,那些尘封的基因便能被随心所欲地激活了。
当然,对于甲藻来说,这还远远不够。众所周知,DNA 具有双螺旋结构,但你或许不知道,这个“螺旋”本身也具有多种形态。目前已发现的具有生物活性的三种螺旋形态分别被称为A 型、B 型和Z 型,其中右旋的 B 型最为常见,而左旋的 Z 型通常只在特定的时期产生。整体看下来,一般的生物体内 Z 型的 DNA 占比都不超过 0.3%。可甲藻显然不是一般的生物,事实上,它只看重了 Z 型结构的一个优点:可以更加紧密地堆积。于是,为了继续压缩,甲藻索性把核内的很大一部分 DNA 转变成了 Z 型。

所以,下次你再碰到有人说地球生物的 DNA 只有右旋,没有左旋时,就让他来看看甲藻。此时他就会知道,什么左旋镜像生命,那都是自然界玩剩下的。(好吧,这是玩笑话;真正的镜像生命概念还要求蛋白质的手性也翻转过来,而甲藻显然还没到这一层)
言归正传,在这一系列工作完成之后,甲藻细胞核的空间利用率已经远远超过其他真核生物了。那么,当你有了这么多剩余的空间后,会用它来干什么呢?当然是用各种诡异的东西把它塞满。在经历了多次丧心病狂的基因组复制和水平基因转移后,甲藻的细胞核中已经是字面意思上的包罗万象了;各种来自动物、植物、真菌、古菌、细菌、原生生物和其他藻类的基因都在此共襄盛举,真是一片勃勃生机、万物竞发的境界。
到了这个份上,甲藻核似乎已经成为了完全体,没有什么可以再改的地方了。今天的夜光藻纲(Noctilucophyceae)就大致保留了这样的细胞核;它们还把光合质体给退化了,并额外演化出了一条触手,从而转变成了浮游掠食者。纲下最知名的物种就是夜光藻(Noctiluca scintillans),拥有直径可达两毫米的巨大细胞以及生物发光能力,从而引发俗称为“蓝眼泪”的现象。
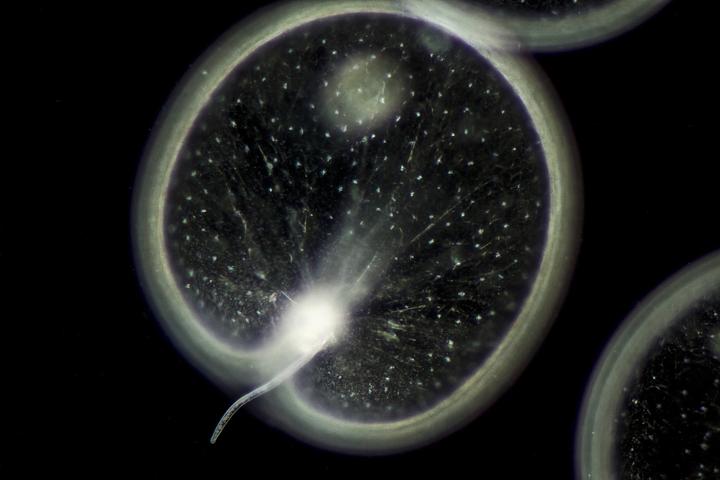

但是,真正的甲藻,也就是甲藻纲(Dinophyceae)表示,它们的细胞核还能继续压缩。于是,它们不知通过什么方法,把 DNA 直接干成了一种超浓缩的液晶态,这样一来就不需要那么多奇奇怪怪的蛋白质拼命压住基因不让它反弹了。毕竟,蛋白质虽然相比 DNA 链来说占据的体积很小,但现在既然其他地方都已经优化到极致了,那也就只能委屈一下它们了。至于有些关乎最基本的新陈代谢的基因由于任务繁重,会专门进行局部的解压,形成拱形基因丝来满足日常所需。
如果你觉得这就是甲藻的极限了,那显然还是太小看它了。在此基础上,甲藻纲的核心类群还将核膜内陷,构建出了一套极为复杂的核内内膜系统(即所谓的核网(nuclear net))。以多沟藻属(Polykrikos)为例,它们的核网主体部分由六条平行的大通道构成,周围有众多的小腔室和小通道将整个系统连为一体:
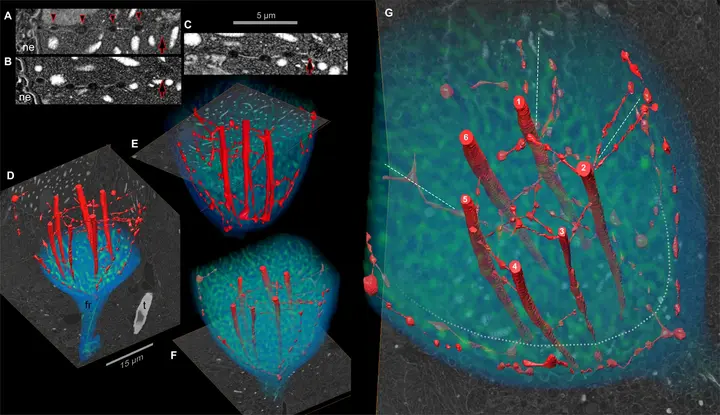
由于核网只在有丝分裂期才会完整的展现,所以这一系统显然与甲藻的有丝分裂密切相关;不过具体怎么个相关法,还需要进一步的研究。
总之,在加上液晶态 DNA 和核内内膜系统之后,甲藻才终于算是彻底完成了细胞核的革命。虽然这一系列变化只是将一座屎山变成了另一座更大的屎山,但你也不得不承认,大屎山确实给甲藻带来了众多生存优势。从三叠纪到侏罗纪,甲藻一路辐射演化,最终成为了海洋光合浮游生物中第二繁盛的类群,直至今天。
况且,身怀一大坨基因的它们压根就不想只局限于光合生物的生态位,而是各种左右横跳。其中最简单的操作就是像夜光藻那样,把质体扔了变成异养生物。著名的鳍藻目(Dinophysales)就是这样,由一个丢失光合能力的祖先演化而来;它们还额外配置了两块超大的甲片,突出细胞本体宛若鱼鳍。关于该结构的功能一直以来都争议很大,但最近的一些研究表明,这个复杂的“鳍”似乎是用来养殖细菌的。具体来说,“鳍”中有很多大大小小的沟壑,内部的微环境很适合某些细菌的生存和繁衍;而当鳍藻感觉饿了的时候,就会从“鳍”中抠一些细菌出来果腹。

如果要邪门一点,也可以请出刚才已经提到过的多沟藻属。它们的细胞在分裂后不会马上分开,而是一个摞一个地形成合胞体,堪称人体蜈蚣。凭借这招,它们硬是凑出了比较大的体型,从而得以逐梦掠食圈:
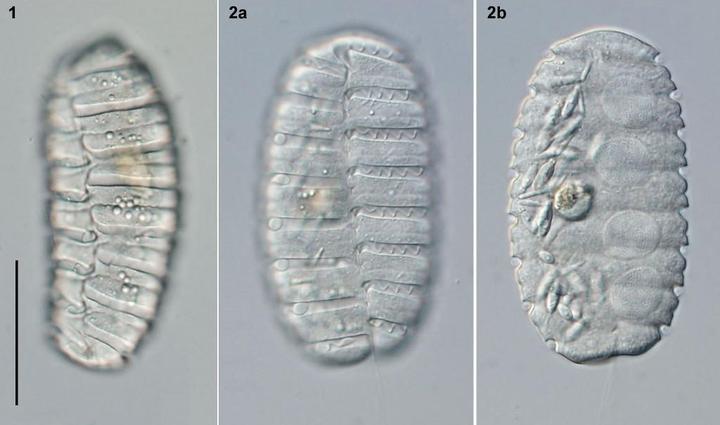
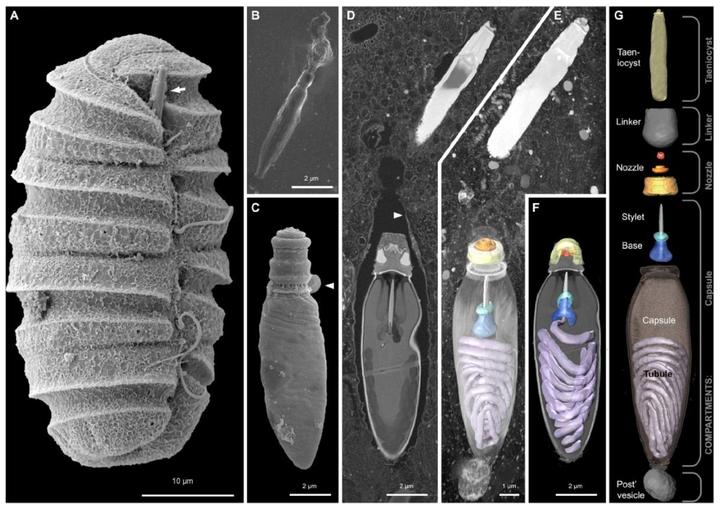
可是,仅靠体型还不够,它们需要一个好用的攻击武器;于是,它们想到了甲藻的另一件神装——刺丝泡(trichocyst)。简单来说,这是一种特化的囊泡,内部包含有一根被压弯的细丝;当有敌人来犯时,这根丝可以高速弹出并扎穿敌人的细胞膜。本来是一个被动防御的结构,但多沟藻将其口径加粗,射程拉长,改造成了一根可以主动射出的鱼叉,用于攻击和固定猎物:
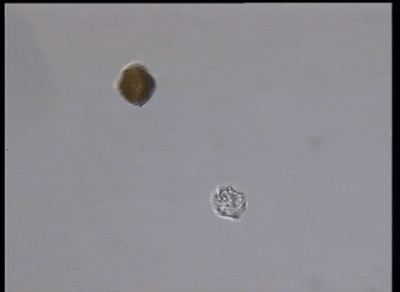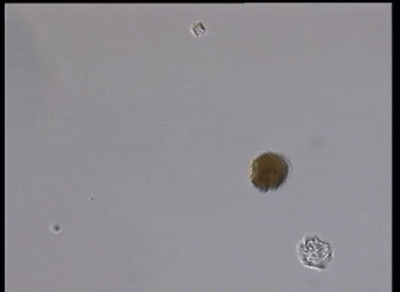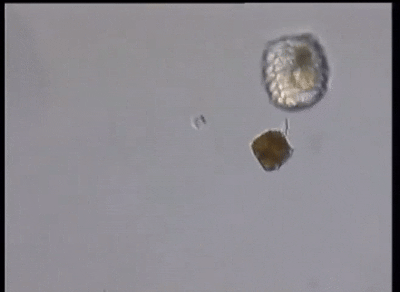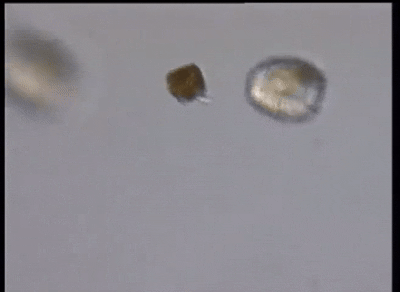
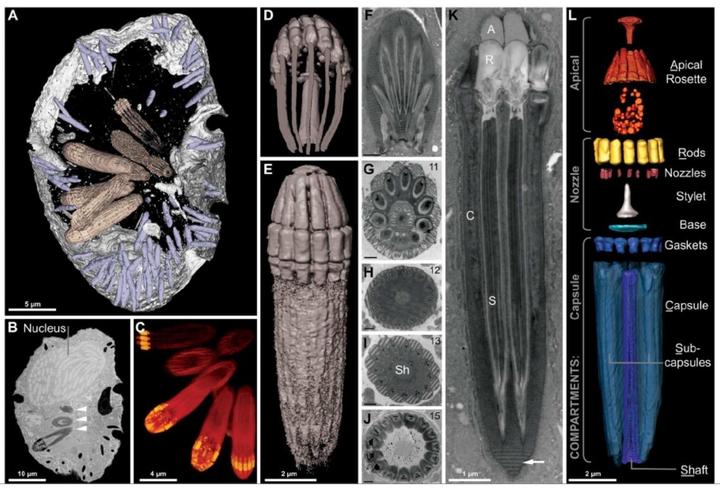
当然,这还远远不是甲藻的战力极限。在掠食这条路上走得最远的,是与多沟藻同属裸甲藻目的单眼藻科(Warnowiaceae)。为了感知猎物的方位,它们以一个单细胞之躯,演化出了一个字面意义上的、完整的照相机眼。这个眼睛主要由三部分组成,外侧的角膜由线粒体爆改而来,中间是负责汇聚光线的晶状体,而内侧的视网膜本质上是一种特化的、失去光合能力的质体,负载有视黄醛和不知道哪天从古菌那里捡来的紫膜质充当感光蛋白;从结构上来说和我们动物的眼睛也大差不差,只不过我们用了几亿个细胞来组装它,而单眼藻只用了一个细胞的一部分:
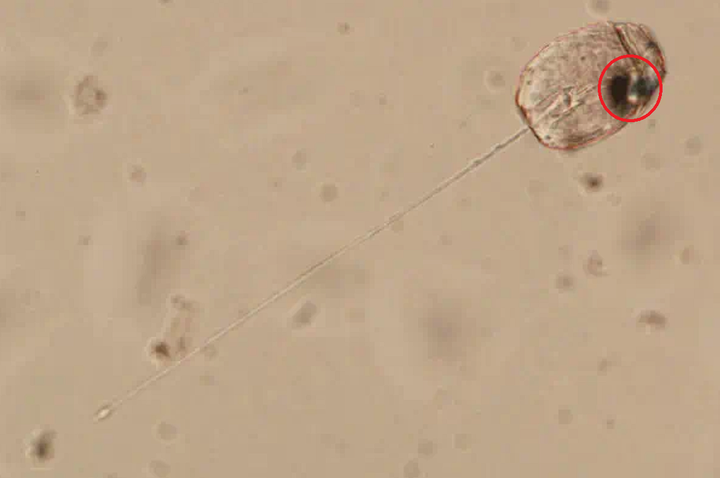
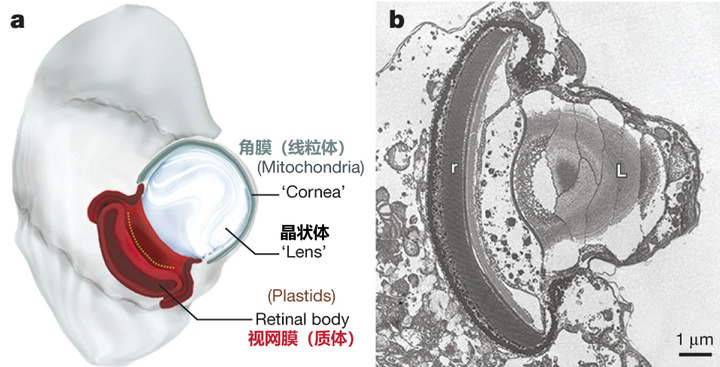
在单眼藻刚被发现那会,人们怎么也无法相信单细胞生物可以演化出眼睛,以至于出现了一种极为邪门的假说,认为这其实是多细胞生物的眼睛恰好被单细胞生物吃掉了,且还没来得及消化。即使是现在,这只眼睛上还有众多的谜团,以至于它是否能够真正成像都无定论;当然,有一点是确定的,那就是这只眼是已知最复杂的亚细胞结构,或许都没有之一。
只不过,对于单眼藻来说,如此逆天的眼睛也只是它满身神装的一部分。科内以单眼藻属(Warnowia)为代表的分支将多个刺丝泡拼到了一起,再结合自身分泌的毒素,组装成一把物法双修的“加特林”。一旦眼睛锁定了猎物,它们就会用这把武器远程射击,而且由于单眼藻都习惯随身佩戴好几把“加特林”,所以攻击时绝对量大管饱:
而以红沟藻属(Erythropsidinium)为代表的分支则没有这么极端。它们在细胞尾部配备了一个可以伸缩的“活塞”,其最大伸长量可达体长的二十多倍,伸缩一次最快仅需三十毫秒,说这玩意是一秒六棍都似乎有点侮辱它;它不仅可以使得红沟藻以一种鬼畜的方式运动,还能够在猎物的防区之外依靠高攻速和魔法伤害(毒素)将其血条即刻清零:


当然,演化为掠食者其实是甲藻向其他生态位进发的过程中,最安分守己的一个选择;而那些不正经的,可就放飞自我了。比如,蛎甲藻属(Ostreopsis)演化出了多细胞群落,而且还放弃了浮游的生活方式,转向底栖;乍一看去,还以为是什么褐藻。更可气的是,这货还很容易在海缸里长,难以去除也就算了,还时不时分泌点剧毒物质,给缸里的鱼虾一点小小的甲藻震撼:

再抽象一点的,比如著名的费氏藻属(Pfiesteria),虽然身为单细胞生物,但可以猎杀比它大得多的多细胞动物。它们会附着在鱼的体表,并直接吞噬鱼的皮肤组织;到一定程度后,它们便会直接钻进鱼的皮肤中大量繁殖,并分泌毒素麻痹鱼的免疫和神经系统,最终往往导致鱼的死亡。另外,如果接触到的是人的皮肤,那费氏藻也不会介意,不过由于其毒素对人类的作用不是很大,所以一般不会致死。
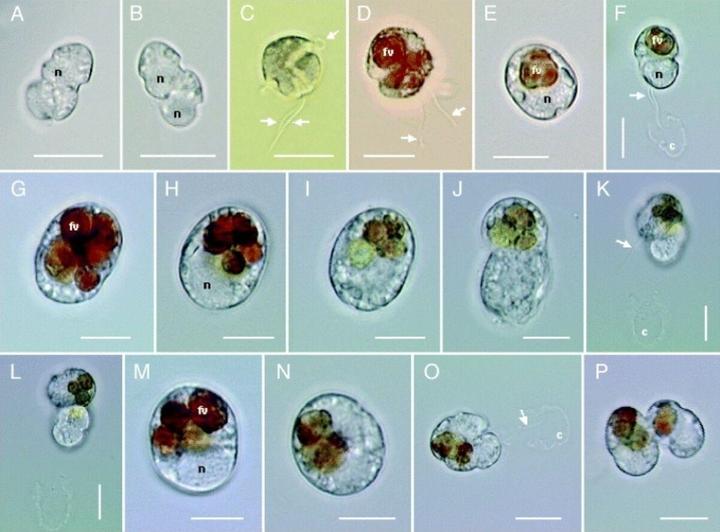
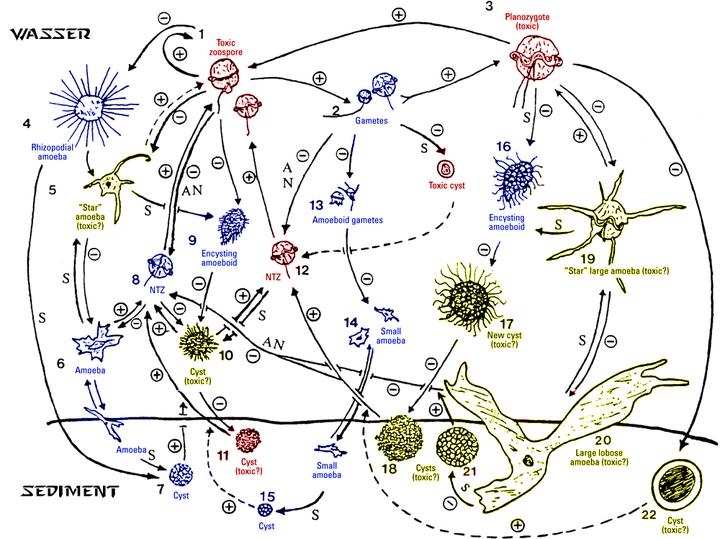
不过,比起这种半寄生性质的越级杀,费氏藻最逆天的地方还是其复杂到离谱的生命周期。在这二十多种不同的形态里,光休眠包囊就有好几种,甚至还有没有鞭毛只有伪足的变形虫形态;这些形态互相之间的转变,更是连变形金刚看了都自愧不如:
当然,在我看来,费氏藻还不是群魔乱舞的甲藻纲中最逆天的,因为在它之上,还有传说中的简虫属(Haplozoon)。乍一眼看上去,你绝对无法相信这玩意是甲藻,因为它长得似乎与某些蠕虫别无二致,包括一个明显的头部(和一个不太明显的尾部)和分节的身体。事实上,早期的生物学家也是这么想的,并将其归为了中生动物(Mesozoa):

只不过,它特殊的甲藻核以及体内退化的质体终究还是出卖了它的身份。事实上,这货的身体结构也和其他几乎一切生物都截然不同。它全身上下只有一个细胞,但被一种特殊的隔膜分成了好几块,每一块都有一个细胞核,相当于一个“半独立”的细胞,而这种形态就是所谓的分隔合胞体。这种身体结构使得简虫可以一定程度上地进行“细胞分化”,比如前端的头部(上图中的 T 部分)拥有一根可以反复射出的中空刺丝泡,是它的进食器;中部的一连串半独立细胞(G 部分)专精无性繁殖,通过分裂把自己的身体变得越来越长,到了一定程度就把其中一个挤出去,成为独立的个体;后端的细胞(Sp 部分)充当孢子囊,用于释放孢子。

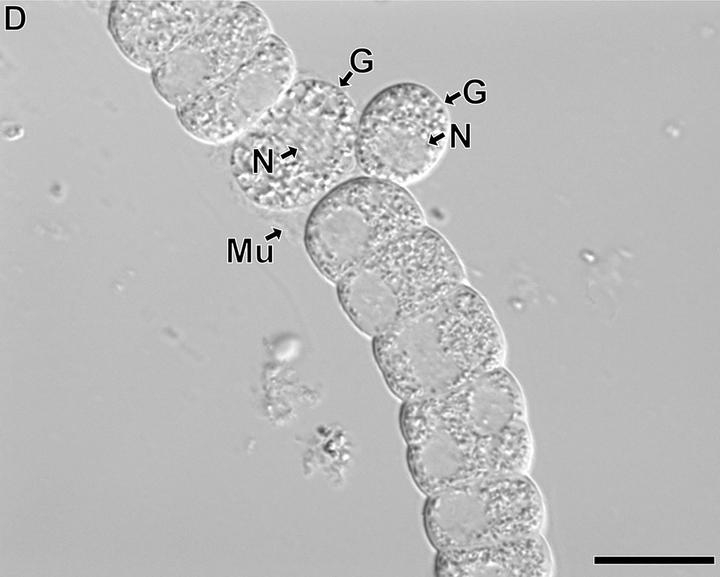
至于它的生活方式,显然是寄生;而且和费氏藻不一样,这货可是纯粹的寄生虫,主要寄生于环节动物体内。
总之,甲藻纲可谓是在各种生态位上遍地开花,而这也多亏了它们那包罗万象的细胞核;虽然是傲视其他所有生物的终极屎山,但其中却包含了演化游戏的整个卡池。也不知道在未来,这个诡异的家族还会涌现出怎样超乎想象的生命形态呢?
一些补充与对上文内容的注解:
(1)虽然上文中一直在说甲藻的细胞核带给了它们怎样的优势,但不得不说,万物都有两面性。终极屎山的代价,就是严重拖累了它们的繁衍,并使得它们对于磷等元素有异乎寻常的高要求(毕竟 DNA 链就是由磷酸基团撑起来的)。或许也是正是这个问题,导致它们自中生代以来就一直是浮游光合藻类的老二,而无法成为真正的王者。
(2)许多资料显示,有一些变形虫拥有异乎寻常的巨大基因组,比如多卓变形虫属的Polychaos dubium,基因组被测出了 670Gb 的大小;但是 21 世纪以来,有不少研究对此都表达了强烈的质疑,并认为这一数值是由于较老的测量方法的缺陷和变形虫细胞结构的特殊性所致。在其他近缘物种中已经有基因组大幅缩水的案例(比如最常见的变形虫之一Amoeba proteus)。因此,在绘制文章的第二张图中没有考虑变形虫。
(3)甲藻在投毒这一块非常出名(虽然事实上大多数甲藻并不会制造毒素),诸如雪卡毒素,刺尾鱼毒素,石房蛤毒素等多种知名的、毒性排名前几的毒素都是由甲藻制造并流入食物链的。也是因此,一旦甲藻爆发形成赤潮,往往会对当地生态造成不小的冲击。
(4)关于红沟藻,它的那个“活塞”到底由什么东西演化而来,据我所知目前仍无法下定论。鬼谷在他关于甲藻的视频中明确提到了该结构由鞭毛演化而来,但我在此处提出质疑:红沟藻的细胞本身就具有两根可见的鞭毛(据这篇论文,两根鞭毛虽然都退化得很短,但依然可见),若该结构真由鞭毛演化而来,那是否说明其祖先拥有更多数量的鞭毛?我认为这对于甲藻来说极不合理;此外,我未能找到支持该结构由鞭毛演化而来的文献。
(5)甲藻核的特殊性,使得对其的分子生物学研究也遭受了巨大的困扰,再加上许多前人根据形态划分的类群没有单系性可言,导致如今甲藻纲内部的分类极为混乱。如果你对这一块感兴趣,或者想进一步了解甲藻及其近缘类群之间的关系,可以看看下面这篇文章: