
人类起源于非洲是没有争议的,有争议的仅是人类究竟多少次走出非洲。目前年代超过两百万年的古人类化石都发现在非洲。非洲有沙棘人、千禧人、南方古猿、肯尼亚平脸人、能人、鲁道夫人、树居人等早期人族的化石,欧亚大陆没有这些化石,欧亚大陆发现的最早的古人类化石就是直立人了。即便是支持多地起源说的学者,对于欧亚大陆直立人来自非洲这点也没有异议,他们与非洲单源说的分歧在于直立人走出非洲以后的世界各地人类的演化过程。欧亚大陆的古人类化石分为三类,第一类是直立人,第二类是古老型智人,第三类是现代人。多地起源说认为世界各地的直立人 - 古老型智人 - 现代人是一脉相承的,不同地区的直立人分别演化成了现代人。非洲单源说则认为全世界的现代人都有着共同的起源,他们的祖先都是非洲智人,现代不同族群是很晚才分化出来的。并认为人类曾多次走出非洲。第一次是直立人,第二次是尼安德特人等古老型智人,最后是现代人。
多地起源说过去曾经非常流行。但是随着分子人类学兴起,DNA 研究证据为非洲单源说提供了强有力的支持。1990 年代以后,赫托长者智人化石的发现与奥莫、杰贝尔依罗等化石的年代测定也为非洲单源说提供了化石证据。化石显示解剖学意义上的现代人在二十万年前就已经出现在东非了,欧亚大陆出土的同时期古人类化石和现代人相差甚远。更重要的是,欧美科学家已经获取了欧洲与西伯利亚古老型智人的 DNA,这些被称作尼安德特人与丹尼索瓦人的古老型智人在遗传上和现代人区别很明显。目前的 DNA 与化石证据支持人类多次走出非洲的说法。
分子人类学为何支持非洲单源说
分子人类学支持单地起源说是因为世界各地现存人群之间的遗传差距很小,他们的分化比想象中要晚,大部分现存人群都是 10 万年内分化出来的。即便是最早分化的科伊桑人,与其他人的分离也只有十几万年。现代人的共同起源地被认为是在非洲,是因为非洲族群与非洲以外族群在基因多样性上并不是均等的,非洲的基因多样性远远高于非洲以外。
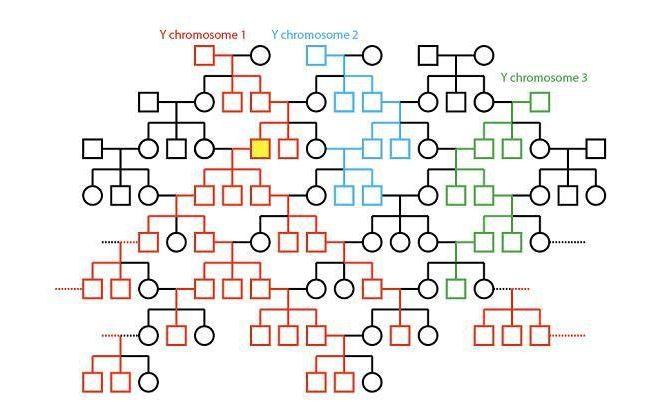
在所有为我们提供 DNA 的祖先中,提供 Y 染色体 DNA 和线粒体 DNA 的祖先最容易追溯。Y 染色体是男性的性染色体,它由父子相传,每个男性的 Y 染色体都来自他父亲的父亲的父亲。线粒体是细胞核之外的细胞器,受精卵的线粒体均来自卵子,因此它由母系传承,所有人的线粒体都遗传自母亲的母亲的母亲。分子生物学支持非洲单源说是从线粒体研究开始的。1987 年美国科学家艾伦·威尔逊、马克·斯通金、丽贝·L·卡恩等人对不同种族女性的胎盘细胞中的线粒体 DNA 进行研究后发现,世界各地的人的线粒体 DNA 差异有限,且非洲人的线粒体 DNA 相对其他人具有更高的多样性。他们发布论文提出所有现代人的线粒体基因都可以追溯到距今大约 20 万年前的一位非洲女性。这位女性被称作线粒体夏娃。到了 90 年代,对人类 Y 染色体的研究也得出了类似结论,全世界男性的 Y 染色体都可以追溯至来自非洲的一位男性。他被称作 Y 染色体亚当。
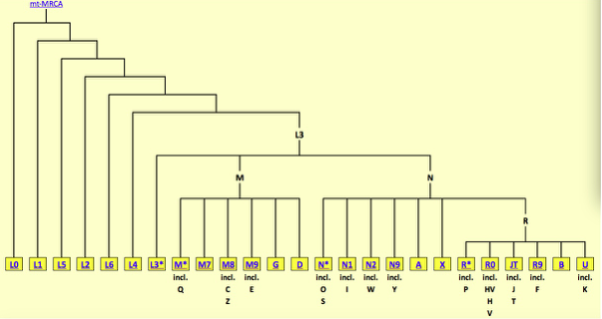
尽管人们的 Y 染色体与线粒体拥有共同的来源,但是从 Y 染色体亚当 / 线粒体夏娃的时代传至今天,在几千代的时间里。他们的后代的 Y 染色体 / 线粒体 DNA 也逐渐出现了不同的突变,从而形成了不同的单倍群。分子人类学家为这些单倍群分别命名并建立了树状的 Y 染色体 DNA 谱系与线粒体 DNA 的谱系。线粒体 DNA 谱系树中最早分化的支系都分布在非洲,最先分化的是 L0 与 L1,接下来依次是 L5、L2、L6、L4,然后是 L3,最后 L3 分化出了非洲以外的两大超级祖母——M 与 N。所有非洲以外的人都是 M 或 N 的后代。虽然非洲的单倍群均以 L 命名,但这些 L 之间其实区别很大,它们的多样性远高于非洲以外的单倍群。L4 与 L3 的差异要大于 L3 与任意一个欧亚大陆单倍群之间的差异,那些更早分化的单倍群更是如此。
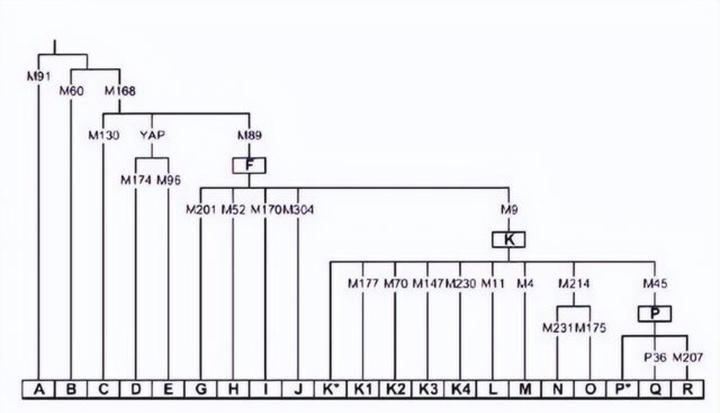
Y 染色体 DNA 谱系树中最早分化的支系也同样都分布在非洲。大部分非洲男性和所有非洲以外的男性的 Y 染色体都带有 M168 突变,他们都是 CT-M168 的后代。较早分化的 A、B 仅分布于非洲。Y-DNA 的 A 群和 mtDNA 中的 L 一样是旁系群而非单倍群,不同的 A 彼此之间的关系并不比他们与其他单倍群的关系更密切。目前发现的最早分化出来的单倍群是 A00,它大约在 27.5 万年前与其他单倍群分化。A00 起初于 2013 年在一名非裔美国人身上被发现。此后,西非多个族群中发现了这一罕见的单倍群的踪迹。2020 年,美国科学家在喀麦隆 Shum Laka 遗址一名 8000 年前男孩的 DNA 中发现了 A00,该男孩的常染色体 DNA 显示他在遗传上介于西非黑人与俾格米人之间。另一个较早分化的支系是 A0,它仅少量分布于俾格米人中。其他不属于 A00 也不属于 A0 的单倍群都是 A1 的下游,A1 包括 A1a 与 A1b,A1b 的下游包括 A1b1 与 BT。BT 是单倍群 B 与 CT 的上游,它实际相当于 A1b2。
由于 Y 染色体亚当与线粒体夏娃顶着亚当夏娃的头衔,所以经常有人问,在他们所处的时代难道没有别的男人和女人吗?其实,他们所处的时代当然有其他的男人和女人,只是他们的 Y 染色体与线粒体没有流传下来。Y 染色体 / 线粒体的传递并不像人们想象中那样稳定,它们必须世世代代都生出同一性别的后代才能传递下去。一旦某一代未能生出指定性别的后代,这一支就绝嗣了。这是不可逆的,假设某一女人的某一代后人生出的全是男性,她的线粒体就没了,即便这些男性后代下一代生的全是女孩,她们的线粒体也不是继承自奶奶。在这种机制下,Y 染色体与线粒体的淘汰率是很高的。可以参考东亚人有没有可追溯的超级祖先? 东亚的主要 Y-DNA 祖先,年代超过 3100 年的有几千个,7300 年前的就只有几百个了,超过 16500 年的更是只有几十个,绝大部分东亚男性的 Y 染色体都来自这几十个男性。这几十位超级祖父的后代世世代代都生了足够多的男孩,与他们同时代的绝大部分男性都没有做到这点。人们的线粒体祖先也同样是越往前越少的,超级祖母们的后代世世代代都生了足够多的女儿,与她们同时代的绝大部分女人都没有做到这点。一些未能将 Y 染色体与线粒体流传下来的男人和女人同样是我们的祖先,他们是我们的外公与奶奶。他们虽然未将 Y 染色体与线粒体流传下来,却在常染色体 DNA 方面对后代有所贡献。
相比 Y 染色体或线粒体,常染色体 DNA 更加重要。如果将人们的 DNA 比作一本书,Y-DNA 与 mtDNA 就只是其中两页。常染 DNA 是剩下的内容,它决定着人们的体貌特征与其他遗传性状。和 Y 染色体或线粒体一样,常染也会出现基因突变的现象,比如东亚人与美洲原住民都普遍拥有 EDAR-V370A 衍生等位基因,它带给了我们粗而直的头发,更密集的小汗腺,还与铲形门齿相关。这个基因来自三万年前的一位祖先,他生活的年代,美洲原住民还没有和东亚人分离,所以我们共享这个基因。当时已经与东亚人分道扬镳的欧洲人、非洲人则缺乏这个基因。在每个族群中都有许多像 EDAR-V370A 这样的衍生等位基因。它们在不同族群中的频率差异造就了不同人群的种族特征。
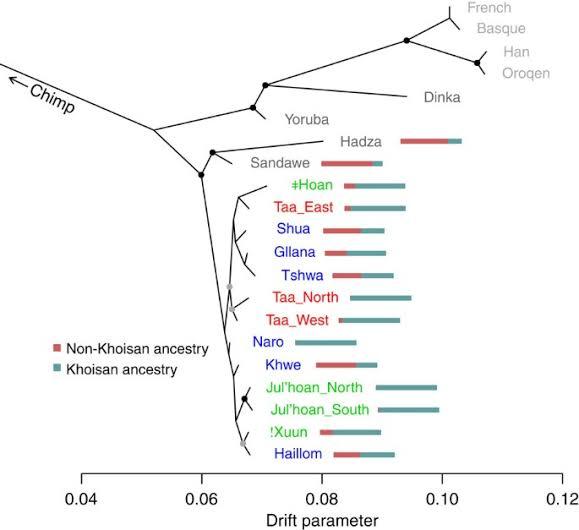
尽管非洲以外的一些族群会因种种原因认为自己很独特,但基于常染色体 DNA 的研究显示,非洲人的常染基因多样性同样远高于非洲之外。在所有现存族群中,与其他人遗传距离最大的是南非原住民科伊桑人。[1]他们在十几万年前就已经和其他人分化了,有些研究甚至认为他们与其他人分化的时间可达 20 万年。其次是中非原住民俾格米人,一些研究认为他们可能和科伊桑人同样古老。[2]其余人的分化时间都很晚,包括走出非洲的欧亚人群与占如今非洲人口大多数的西非黑人、东非黑人、班图黑人等主流黑人族群。其中西非黑人与欧亚人群(及东非黑人)分化的时间稍早,约在 7.5 万前。东非黑人与欧亚人群分化的时间稍晚,约在 6.5 万年前。[3][4]
这是根据科伊桑人各部族与约鲁巴人(西非尼日尔 - 刚果语族)、丁卡人(东非尼罗特人)、欧洲人、东亚人的遗传距离构建的邻接树。这些来自非洲、亚洲与欧洲的人群可以明显的分为两支,科伊桑人与非科伊桑人。就像非科伊桑人分为很多支系一样,科伊桑人内部也有不同的支系。不同支系的科伊桑人之间的遗传距离就像欧洲人与东亚人的差异一样大。[1]
这是纳米比亚 Nama 文化节上的 Nama 人,Nama 人是科伊科伊人的一支。科伊桑人分为科伊科伊人与桑人两大分支。科伊科伊人和桑人都有着黄褐色的皮肤,胡椒粒状卷发,内眦褶出现率较高。但他们也有区别,科伊科伊人的肤色更浅,脸更平,五官较小巧。

已故影星历苏来自桑人部落,他们是科伊桑人的另一个分支。

除了科伊桑人、俾格米人以外,非洲古代可能还曾经存在过一个更早分化的现代人分支。这是西方学者推测的非洲上古族群分布。地图上红色的代表科伊桑人,他们当时生活在非洲南部。黑色的代表俾格米人,他们当时生活在非洲中部。蓝色的代表一个已不存在的族群,他们可能生活在撒哈拉地带。现代西非人有大约 10%的血统来自这个族群。[2]
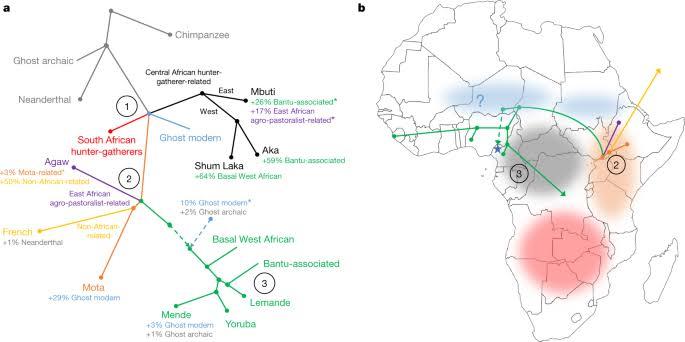
非洲主流黑人与现代欧亚人群的共祖起初生活在东非。12 万 -7.5 万年前,生活在非洲南部的科伊桑人曾大规模向东非扩张。[5]之后东非的人群开始向全世界迁徙,他们中的一支向西迁徙至西非,成为了如今西非黑人的祖先。另一支离开了非洲,成为了如今非洲以外人群的祖先。剩下的留在了东非,他们是如今生活在东非北部的尼罗特人与埃塞俄比亚人。此后,黑人与科伊桑人、俾格米人相安无事的在非洲生活了几万年。直到四千年前,随着农业发展,西非人口大幅度增长,西非黑人中的班图语族群开始向东向南扩张。以狩猎采集为生的科伊桑人、俾格米人完全不是他们的对手。所以班图人逐渐成为了非洲赤道以南地区的主流人群,科伊桑人、俾格米人则变成了少数群体。
化石证据也支持非洲单源说
非洲单源说不仅有分子人类学的支持,它也有古人类化石为依据。迄今为止,世界上发现的最古老的晚期智人化石是出土于埃塞俄比亚南部奥莫基比什(Omo kibish)的奥莫 1(OmoⅠ)。新修复的髋骨显示她应该是一名女性。[6]她之前被认为生活于 19.5 万年前,但最新的研究显示她的生活年代至少在 23 万年以前。[7][8]在同时期的人类化石中,奥莫 1 非常引人注目,她有着饱满的球型颅顶,微翘的下巴,眉嵴也不像其他古人类那样显著。她是解剖学意义上的现代人。
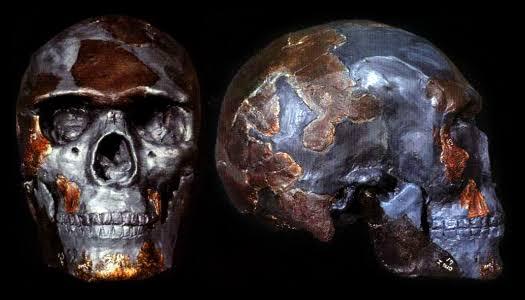
在非洲还有比奥莫 1 更古老的智人化石,其中最古老的是北非摩洛哥距今 31.5 万年的杰贝尔依罗(Jebel Irhoud)人。这是杰贝尔依罗人与尼安德特人及现代人的比较。a 是西班牙 Sima de los Huesos 距今 43 万年的早期尼安德特人头骨。b 是法国 La Ferrassie 距今 6 万年的晚期尼安德特人头骨,c 是杰贝尔依罗人头骨。d 是法国 Abri Pataud 距今 2 万年的晚期智人头骨。[9]我们可以看到相比尼安德特人,杰贝尔依罗人明显更接近现代人。
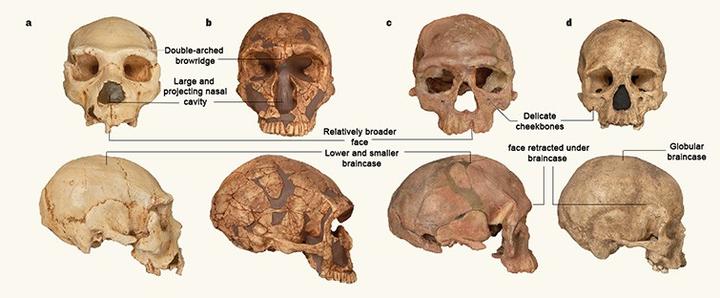
但杰贝尔依罗人与奥莫 1 不同,因为他们尚未发展出现代人的一些关键特征。杰贝尔依罗人的脑容量(1375-1467ml)与现代人相仿,但颅高明显低于现代人,他们的颅腔和现代人存在差异。[10]奥莫 1 则和现代人有着相同的颅腔。杰贝尔依罗人也不像奥莫 1 那样拥有突出的下巴。杰贝尔依罗人刚被发现时曾被误认为是尼安德特人与现代人之间的过渡类型或混血儿,直到他们的年代被测定为 31 万年前,他们才被视为现代人的祖先。而奥莫 1 从被发现的时候起,就是没有争议的晚期智人。
杰贝尔依罗人的年代比奥莫 1 更早,奥莫 1 比他们更接近现代人是情理之中的事。但即便是在同时代古人类化石中,奥莫 1 也很引人注目。除了奥莫 1 以外,埃塞俄比亚还有另外一些与她年代接近的著名古人类颅骨化石,包括同样出土于奥莫基比什的奥莫 2(Omo Ⅱ,距今 23 万年)和出土自埃塞俄比亚北部阿法尔州的长者智人 Herto man(距今 16 万年)。奥莫 1、奥莫 2 和 Herto man 都比杰贝尔依罗人更接近现代人,但他们之间也有区别。其中奥莫 1 比其他人更接近现代人,她的头骨较薄,颅高最高,眉骨最不显著。相比之下,奥莫 2 和现代人差异较明显。Herto man 则介于两者之间。有些研究者怀疑这些出土自同一地区的古人类存在种族差异,认为奥莫 1 和奥莫 2 可能分别来自不同族群。也有人认为他们来自同一个族群,只是该族群存在较大的个体差异。这一时期的东非智人整体上还与现代人存在差异,但他们中的一些人已经与现代人没有多少区别了。

现代人从出现在非洲到走出非洲中间隔了很长一段时间。在非洲以外发现的最古老的现代人化石来自以色列 Skhul 与 Qafzeh 遗址,距今约 10-12 万年。Skhul-Qafzeh 人除了颌部稍突出,其他地方与现代人差异不大。他们平均身高 1.77 米,有着瘦高的身材和修长的四肢。[11]这与尼安德特人不同,尼安德特人身材壮实,四肢相对偏短。这些智人化石出土于 1934 年,由于一同出土的还有多具尼安德特人化石,他们起初曾被当成尼安德特人向现代人过渡的证据。但 1980 年代更精确的年代测定显示,他们的生活年代比一起出土的尼安德特人要早三万年左右。Skhul-Qafzeh 人可能代表着一次失败的走出非洲。他们在 10 万年前居住在黎凡特地区。大约 7.5 万前,地球上出现了剧烈的环境变化,西亚的气候变得寒冷干燥。Skhul-Qafzeh 人消失或离开了这个地区,尼安德特人取代他们成为了这些洞穴的新住户。[12]
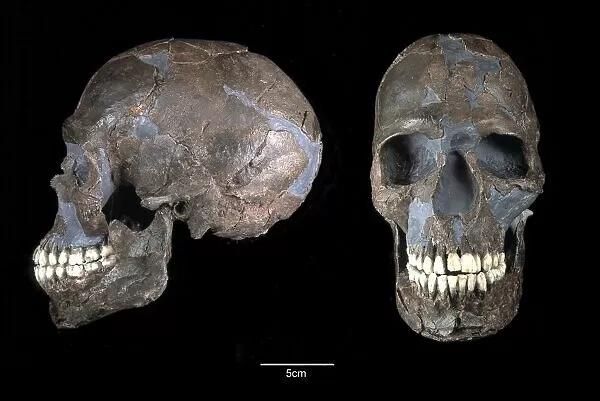
欧亚大陆出土的绝大部分晚期智人遗骸年代都在 7 万年以内,他们是典型的现代人,与分子人类学中 6.5 万年前走出非洲的人群对应。包括欧洲的克罗马侬人和中国的柳江人、山顶洞人等。这些人有许多共同的特征。比如他们都有着低矮的眼眶,较小的上面高,较阔的鼻型(但古欧洲人普遍鼻梁非常高),长颅型,眉骨略明显。[13]针对古欧洲人的基因研究显示,他们还有深色的皮肤。[14]这些都是走出非洲时的晚期智人祖型。虽然智人来自非洲,但智人祖型并没有格外接近现代非洲人。2007 年,F. E.Grine等人对比了南非出土的 3.6 万年前的晚期智人 Hofmeyr Man 与同时代欧洲晚期智人及各地现代人的颅骨形态。发现 Hofmeyr Man 形态上最接近同时代的欧亚大陆晚期智人,而与现代各族群均存在差异。在现代各族群中,大洋洲原住民族(而不是非洲人)相对较接近这些古人。[15]
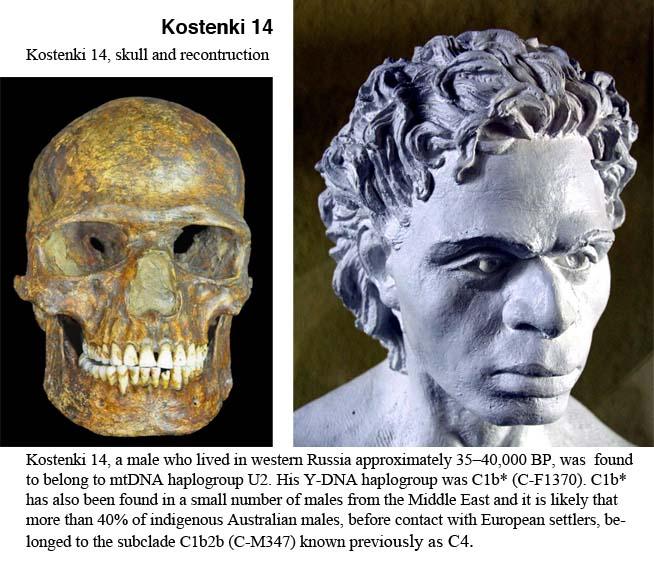
这是 Kostenki 14 复原像。Kostenki 14 出土于俄罗斯西部,他生活于 3.8 万年前,是最古老的欧洲晚期智人之一。他具有高鼻深目的特征,但也有一些异于现代欧洲人的地方。比如他的鼻型较阔,齿槽突颌比现代欧洲人稍明显。2014 年,Seguin Orlando 等人提取了 Kostenki 14 的 DNA 并对其进行了全基因组分析。研究结果显示,Kostenki 14 在遗传上接近如今欧亚大陆西部居民。研究同时还显示他有着深肤色与黑眼睛。[16]
Sungir 遗址古人同样来自俄罗斯西部,是三万年前的古欧洲人,遗传上均属于西欧亚人群。

一万年前的英国切达人(cheddar man),是西欧狩猎采集者的一员。他具有很多现代欧洲人的特征,比如蓝色的眼睛,但他的皮肤是深色的。[14]

东亚晚期智人的代表是山顶洞人与柳江人,由于出土地层不确定,这两个化石的年代都存在争议。山顶洞人的年龄一般认为在 1 万至 3.3 万年之间。柳江人的年龄争议更大,我们只能说他的年代至少在 1.5 万年以上,可能在 6 万年以上。[17]山顶洞人和柳江人与现代东亚人有一些区别。他们有着低矮的眼眶,眉骨比现代东亚人明显,颅骨较狭长,鼻子宽阔。这些都是他们那个时代的智人共同拥有的特征,却不是现代东亚人的特征。[18][13]山顶洞人的头骨在二战时已丢失,现在只有复制品存世,因此无法提取他们的 DNA。不过,同样出土于北京周口店且年代比山顶洞人更早的田园洞人进行过基因测序,他在遗传上接近欧亚大陆东部各族群与美洲原住民,而非欧亚大陆西部人群或澳大利亚原住民。[19]柳江人颅骨比山顶洞人接近现代中国人,他应该也属于东欧亚人群。[20]
这是山顶洞人头骨。其中最左边的山顶洞 102 头骨存在人工变形的情况,她幼年时曾长期缠头拉长头骨,这种故意拉长头骨的习俗和玛雅人有些相似。

另一个著名的东亚晚期智人是日本的港川人。港川人出土于日本冲绳县,他的年代没有争议,可以确定在 1.6-2 万年前。港川人在遗传上最接近绳文人,不过他不是绳文人的直系祖先,而是旁系祖先。绳文人与港川人都属于东欧亚人群。[21]

与奥莫 1 同时代生活于东亚的古人类与非洲智人有很大的区别。这个化石出土于哈尔滨,被命名为龙人,他生活于 14.6 万年 -30.9 年前,研究他的倪喜军、季强等人认为他与同时代生活于中国境内的大荔人、华龙洞人、金牛山人、夏河人都有密切的亲缘关系。[22]龙人的头颅非常大,脑容量则和现代人相仿(1420ml)。他的眉嵴很突出,额头低平后倾,和现代人差异很大。许多西方学者认为他属于丹尼索瓦人的一个支系。[23]丹尼索瓦人是分布于亚洲东部的一支神秘古人类。科学家起初通过俄罗斯阿尔泰山脉丹尼索瓦洞中残存的牙齿与指骨获取了他们的 DNA 数据,得知他们和尼安德特人具有亲缘关系,两者大约在 38 万 -47 万年前分化。[24]中国青藏高原东北部曾出土过一块带臼齿的古人类下颌骨,被称作夏河人。此前的古蛋白质组学研究显示他的牙齿蛋白序列很接近丹尼索瓦人,在其出土洞穴的沉积物中还找到了丹尼索瓦人的线粒体 DNA。[25][26]龙人仅存的一颗臼齿很接近夏河人,倪喜军等人的论文中认为夏河人与龙人的关系甚至比金牛山人更密切。龙人可能属于丹尼索瓦人不仅是西方学者的看法,倪喜军等人在论文中也承认存在这种可能性,只是他们认为现在还不能下结论。[22]
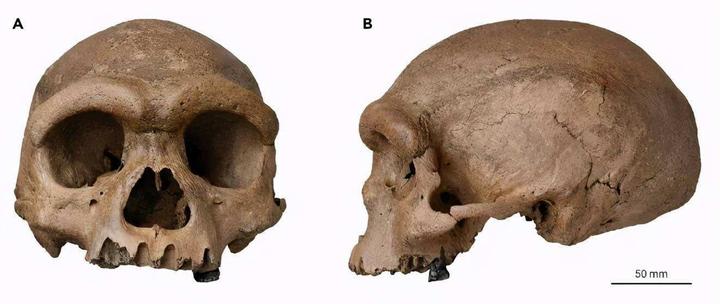
欧洲古人类也同样和非洲智人有很大的区别。这个化石名为费拉西人(La Ferrassie man)[27],是 6 万年前生活于法国的欧洲尼安德特人。尼安德特人曾广泛分布于欧亚大陆西部的欧洲、西亚、中亚等地,他们在那里一直繁衍生息到 3-4 万年前,智人走出非洲后曾与他们共同生活过一段时间。尼安德特人是被研究最多的一支古人类,他们与非洲智人具有同源关系,大约在 55 万 -75 万年前与非洲智人分化。[24]现在多数科学家认为尼安德特人和现代人是同一物种(智人)的不同亚种。他和上面的龙人一样,具有头颅硕大、眉嵴突出、额头低平后倾等特点。尼安德特人的颌部不是很突出,但他们没有现代人那种明显的下颏,这也是他们和现代人的一个区别。
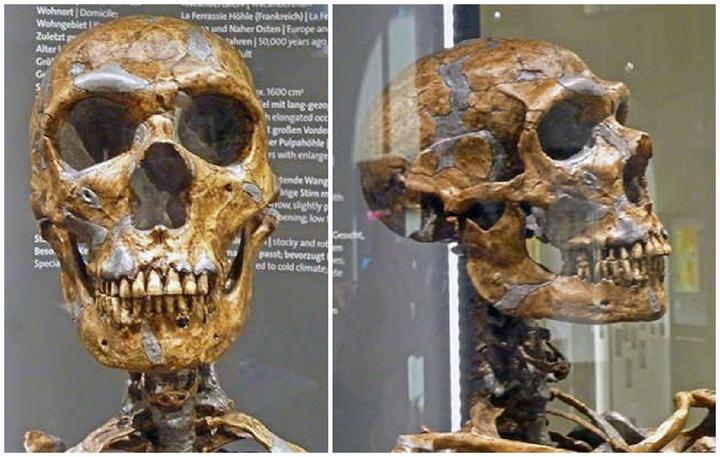
尼安德特人的额头比智人小,脑容量却不低,实际上一些尼安德特人化石的脑容量非常大。比如费拉西人的脑容量为 1643 ml。出土于以色列的中东尼安德特人 Amud 1 号脑容量则高达 1747 ml。[10]但他们中也有一些人的脑容量并不那么大,比如 7 万年前生活在南欧的女性尼安德特人直布罗陀 (Gibraltar)1 号脑容量约为 1213 ml。总的来说,他们的脑容量处于现代人的变异范围内,其平均值略高于现代人的平均值。
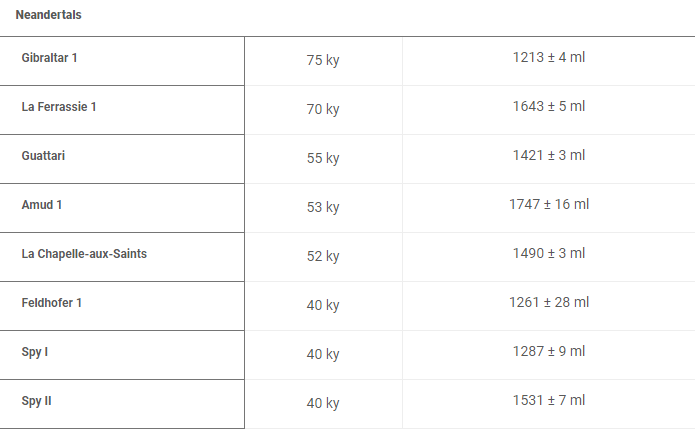
尼安德特人的发育模式和现代人相仿,幼童会吃奶到两岁左右。[28]在七、八岁的时候换牙。[29]尼安德特人儿童有饱满的额头,眉嵴也并不突出。幼年的尼安德特人虽然和同年龄的智人存在差异,但这种差异相比成年时期的差异要小。尼安德特人与现代人的颅型差异是随着年龄增长越来越大的。
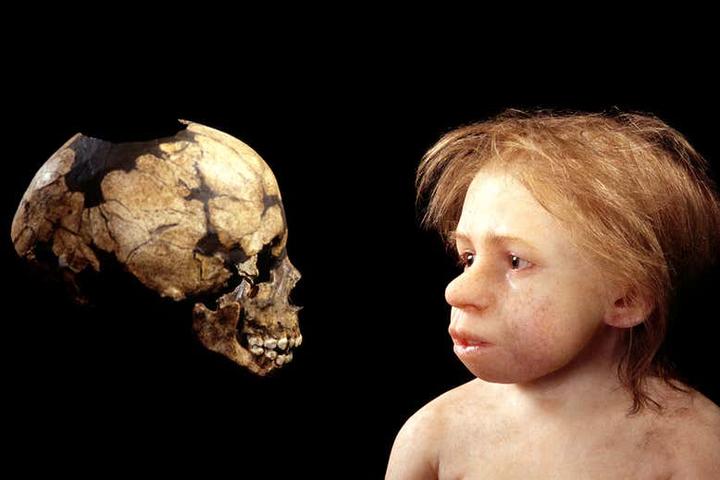
除颅型差异以外,尼安德特人的身材比例也与现代人存在差异。他们的头颅更大,身材更粗壮。有更大的肺活量和更强的力量,特别是手掌的力量远远高于现代人。
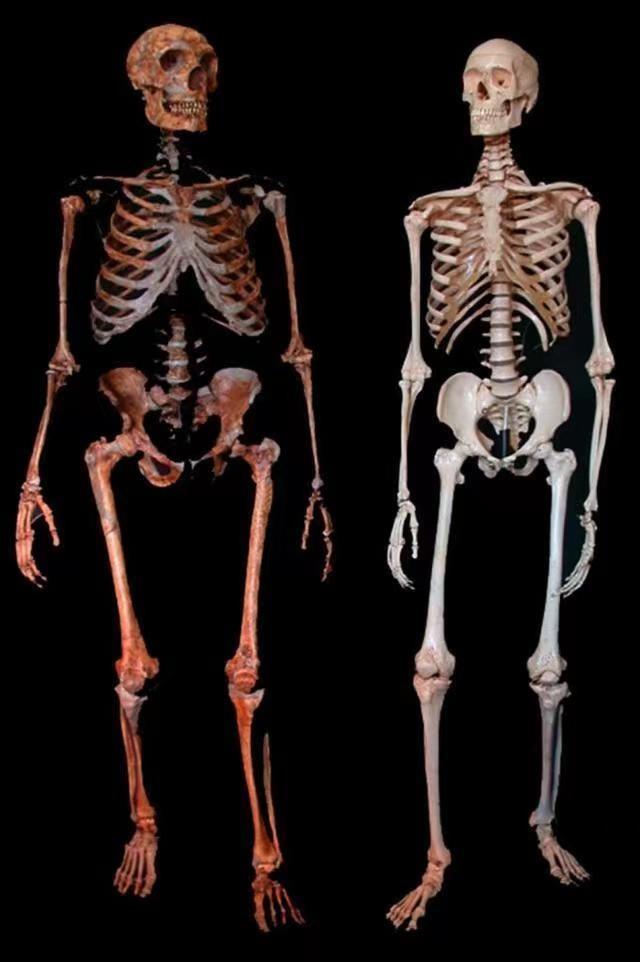
尼安德特人也有一些能让非洲以外的现代人感到亲切的特征。比如生活在欧洲的尼安德特人有白皙的皮肤,还有多种不同的发色与瞳色。2007 年西班牙科学家 Carles Fox 在两名尼安德特人的基因组中发现了与红头发和苍白肤色相关的基因。[30][31]后来另一位科学家又发现另外两个尼安德特人可能具有棕色的头发和棕眼睛。Carles Fox 认为尼安德特人可能有多种不同的发色。[32]虽然尼安德特人的肤色发色和现代欧洲人相似,但欧洲人并没有从尼安德特人那里获得白皮肤与红发基因,他们是趋同演化出来的。[33]

尼安德特人还普遍拥有铲形门齿。[34]不过尼安德特人的铲形门齿形态与现代东亚人并不一样。详情见
尽管尼安德特人和丹尼索瓦人与现代人有很大的差异,但现代人也有少量来自他们的血统。科学家们提取了尼安德特人与丹尼索瓦人的 DNA 与现代人进行对比,发现非洲以外的所有现代人(包括欧洲、亚洲、大洋洲和美洲的居民)都有少量尼安德特人基因。其比例平均在 1.8-2.6%之间。此外,澳大利亚原住民和美拉尼西亚人还有 3-6%丹尼索瓦人基因。东亚人与美洲原住民的丹尼索瓦人基因则在 0.2%左右。[24][35]
在古人类研究领域,过去曾流行将古人类分为不同阶段的阶段论。比如中国学者将欧亚大陆的古人类分为四个阶段,早期猿人、晚期猿人、早期智人、晚期智人。尼安德特人被认为是早期智人。不过,这种划分方式现在已经逐渐被放弃了。尼安德特人不是现代人的直系祖先,更不是直立人演化成现代人的中间阶段。他们和非洲智人是平行的两个不同的支系,有不同的演化路径。目前没有任何证据显示尼安德特人的智力落后于现代人。尼安德特人和现代人一样具有与语言相关的 FOXP2 基因突变。欧洲出土过不少尼安德特人制作的工具,他们会用桦树皮制作沥青,并将沥青当做胶水黏接石矛与木柄。[36]西班牙拉帕西加洞穴(La Pasiega)还出土了距今 6.5 万年的岩画。那个时候智人尚未抵达当地,所以这些壁画被认为是尼安德特人的作品。他们可能还存在原始宗教信仰。


非洲智人有何过人之处?
关于尼安德特人为何会被智人取代,有很多种假说。不过大部分假说都没有足够的证据证实。真正可以确定的是,尼安德特人、丹尼索瓦人与非洲智人的人口规模并不是相等的。
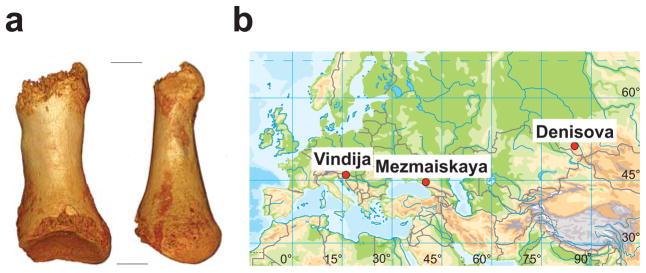
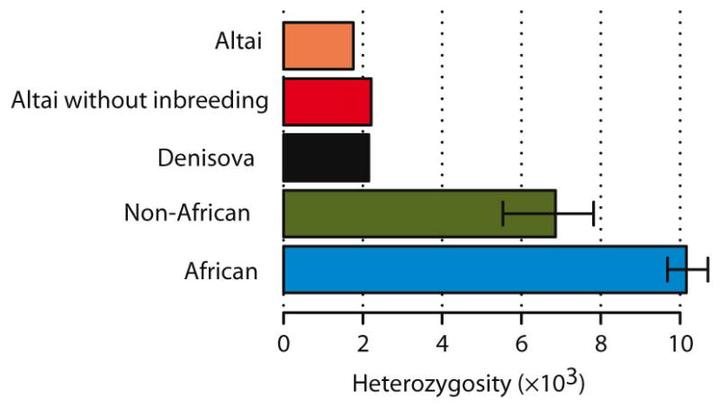
2014 年由多国顶级科学家组成的联合团队对俄罗斯阿尔泰山脉出土的尼安德特人与丹尼索瓦人进行的研究显示。尼安德特人与丹尼索瓦人可能生活于封闭的小群体中。在他们的基因中出现的杂合子位点比例极低。仅相当于欧亚现代人基因组杂合子位点数量的 22–30%,非洲人的 16-18%。这不仅低于现代人,也比世界上大多数生物都低。[24]针对克罗地亚 Vindija 洞的尼安德特人的基因研究也显示他们的基因多样性很低。[35]
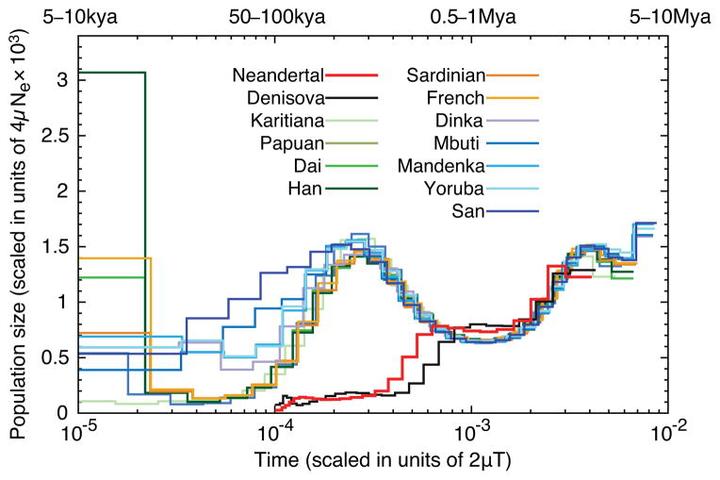
这是通过 DNA 推测的各人群不同时期祖先数量[24],它可以反映人们所属群体在同一地区能够进行基因交流的人口规模。它显示尼安德特人、丹尼索瓦人与现代人的祖先起初是相同的,他们的祖先数量在 100 万年前都曾发生过减少。此后三者走上了不同的道路,现代人的祖先数量重新增长,尼安德特人、丹尼索瓦人的祖先数量则继续减少。尼安德特人分布的范围虽然很广,但他们的人口可能很少。当时的欧亚大陆或许相当地广人稀,以至于同一地区能够进行基因交流的人口非常有限。
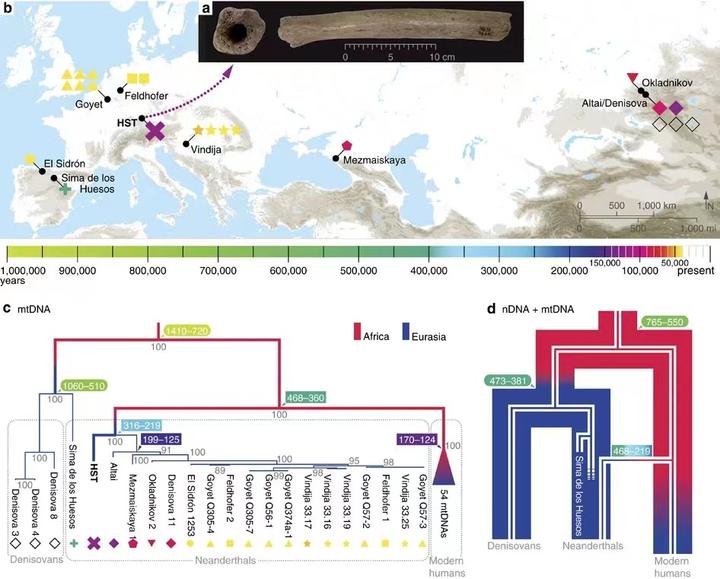
极低的遗传多样性可能会妨碍尼安德特人与丹尼索瓦人及时淘汰有害的等位基因,从而导致遗传负荷增加,这就和近亲结婚对后代的健康有害一个道理。而这有可能造成智人的基因比尼安德特人更具竞争力。2014 年科学家们对阿尔泰山脉丹尼索瓦洞人骨进行研究时发现过一件令人惊讶的事。那就是作为尼安德特人姐妹群的丹尼索瓦人拥有与尼安德特人不同的线粒体 DNA,他们的线粒体大约在 mtDNA 大约在 70–130 万年前与现代人及尼安德特人分化了,而尼安德特人与现代人的线粒体分化时间比这要晚。同时研究还显示尼安德特人与现代非洲人共享的衍生等位基因比丹尼索瓦人要多 7%左右。[24]2015 年 Matthias Meyer 教授从西班牙 Sima de los Huesos 遗址 43 万年前早期尼安德特人的腿骨中提取了线粒体 DNA,研究结果显示他的线粒体 DNA 与丹尼索瓦人相关。2016 年,对 Sima de los Huesos 人 DNA 的进一步研究显示他们的常染接近尼安德特人,而不是丹尼索瓦人。这显示尼安德特人的线粒体 DNA 在后期可能出现了变动。[37]2020 年,针对尼安德特人与丹尼索亚人 Y 染色体 DNA 的研究显示,尼安德特人的 Y 染色体同样比丹尼索瓦人更接近智人。丹尼索瓦人的 Y 染色体大约在 70 万年前与尼安德特人及现代人分化,尼安德特人的 Y 染色体与现代人分化的时间则在 37 万年前。研究者认为比现代人更早走出非洲的智人曾与尼安德特人发生过基因交流,他们的 Y 染色体与线粒体替换了尼安德特人原有的与丹尼索瓦人同源的 Y 染色体与线粒体。[38][39]一个族群只与其他族群进行少量混血,Y 染色体与线粒体就全都被对方替换了,这种事在正常情况下几乎不可能发生。但假如继承智人 Y 染色体与线粒体的个体能够更好的繁衍生息,那么这是有可能的。
这是尼安德特人、丹尼索瓦人、现代人的线粒体与常染 DNA 关系示意图。[40]丹尼索瓦人与尼安德特人在 55-76.5 万年前与非洲智人分离。21.9-46.8 万年前,非洲智人的一支进入了欧亚大陆西部并融入了尼安德特人,他们中的一位女性后来成为了尼安德特人的线粒体夏娃。