





北京周边再次确认华北豹稳定活动,意义不在于多了一条红外影像,而在于它同时证明了三件事:
1.京畿周边山地的生态底盘正在恢复到足以承载顶级捕食者的水平。
2.华北豹正在从核心分布区山西向北扩散。
3.更重要的是,不同区域种群之间出现了重新建立基因交流的现实通道,有望逐步打破长期遗传瓶颈。

要理解这一发现的意义,首先需要回到华北豹本身在生态系统中的角色。
华北豹(Panthera pardus japonensis)是凶猛的伏击型捕食者,适应性强,食谱极为广泛。

从兔形目、啮齿类、鸟类、獾类等中小型动物,到狍子、野猪等有蹄类,均在它的捕食名单上。


通过收集华北豹的粪便并进行 DNA 检测的研究表明,在华北山地森林系统中,狍子(Capreolus pygargus)和野猪是最关键、最稳定的能量来源,二者占据猎物总能量摄入的一半左右。

其种群密度直接决定华北豹的领地规模、活动频率与繁殖潜力。正因如此,近年来关于华北豹的生态研究,往往以豹—狍—野猪这一组合为分析核心。

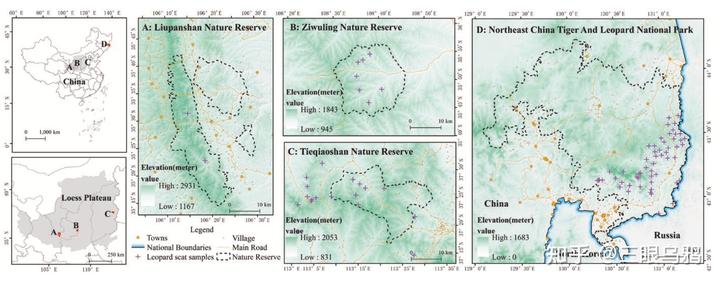
后续的研究,通过在更大空间尺度上系统收集华北豹粪便样本,并引入中国境内另一豹亚种东北豹(亦称远东豹或阿穆尔豹)作为对比参照。

研究人员基于二者的粪便 DNA 宏条形码分析发现:
华北豹共识别 13 种猎物,东北豹识别 15 种猎物,二者共享 8 种猎物,包括狍、野猪、家犬、鼠类等。单个粪样中通常检测到 1–5 种猎物,显示二者都具有明显的机会主义取食特征。
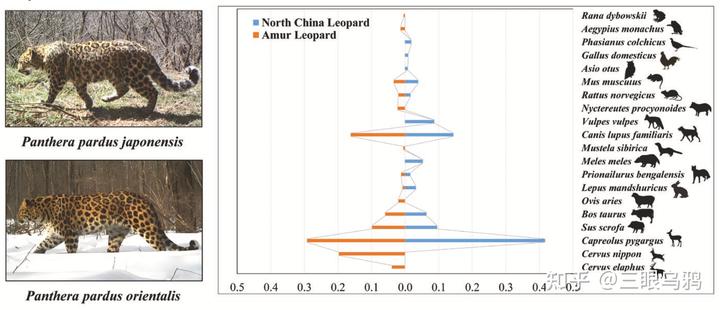
两种豹最常出现的猎物均为狍(华北豹 41.69%,东北豹 29.20%)。此外,家犬在两种豹食谱中的出现均超过 10%,说明人类生活区的影响已深入豹的领域。自由放养犬只进入林区显著提高了与豹的接触概率,不仅加剧人兽冲突,也增加疾病传播风险。

不同的是,东北豹额外从马鹿 *(仅约 3%)和梅花鹿(高达 19.75%)等较大型有蹄类中获取显著生物量。
* 马鹿体重过大,重达 250-300kg,是梅花鹿 2-3 倍。这大大超出了豹的捕猎范围。粪便里出现一般是幼崽或者亚成年,也可能是吃了虎留下的残骸。

而华北豹更依赖中小型食肉动物和兔类(赤狐、狗獾、野兔在华北豹食谱中合计贡献约 18–21%)。黄土高原的破碎化生境可能使中小型猎物相对更易获得。

总体而言,小—中型猎物(<37kg)贡献了两种豹超过 75%的总摄入生物量。

基于出现频率和生物量的分析均显示,两种豹在猎物多样性和生态位宽度上差异不显著,但食性重叠度极高(Pianka 指数约 0.77–0.78),表明其基础捕食策略高度相似,差异主要源于区域猎物群落结构。


需要指出的是,在历史上,华北山地并非只有豹一种大型捕食者。20 世纪上半叶之前,华北地区仍存在虎的边缘分布记录,但至 1950 年代前后,虎已在华北山地彻底消失。在这一背景下,华北豹成为当地唯一仍存的大型食肉动物,在相当多的山地景观中承担了顶级捕食者的功能。

华北豹的捕食行为不仅调控猎物数量(尤其是小型鹿类),也通过改变猎物的取食位置和活动节律,对林下植被更新和群落结构产生连锁影响,因此被广泛视为衡量生态系统完整性的关键指示物种。
栖息地能不能养豹,关键不只在于豹能吃什么,更在于它能不能稳定繁殖并把幼仔带到独立。
基于 2016–2023 年太行山中段长期的红外相机监测数据(覆盖 332 个点位、累计工作日超 41.6 万天),科学家首次揭示了华北豹的繁殖秘密。

监测系统累计记录了 261 次与繁殖相关的行为事件,包括求偶跟随、交配及育幼,确认至少有 57 只成年雌豹参与了繁育,共产生 43 胎,其中近半数为连续繁殖。

更令人欣慰的是,影像捕捉到了 79 只成功独立的幼仔。这意味着,野外出生的幼豹能够顺利度过最脆弱的阶段,完成从出生到短期独立的跨越。

数据显示,华北豹每胎平均成功独立的幼仔约 1.8 只,雌豹首次分娩年龄约 4.6 岁,幼仔在母亲身边生活近 12 个月后走向独立。

这些数据勾勒出一个事实:在太行山的部分区域,华北豹种群已具备了自我更新的能力。
但如果我们把镜头拉到更大的尺度,决定它们长期命运的,仍是核心分布区的总体格局。

数量回升并不自动等于安全。对于大型食肉动物,真正决定未来上限的不是某一地多了几只,而是栖息地是否仍被切割成彼此隔绝的孤岛。
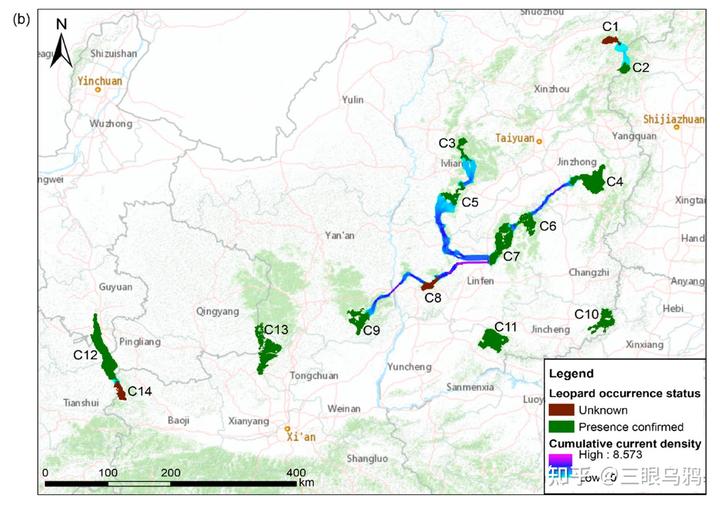
从全国尺度看,华北豹的总体处境仍然脆弱,但在核心分布区已显现出明确的恢复趋势。随着生态保护力度持续加大,华北豹的种群数量在局部区域内稳步回升。

山西省作为目前最重要的核心分布区,长期监测结果显示,可通过个体识别确认的华北豹数量已由 2018 年的约 132 只增加至 2025 年前后的 220 余只。

基于监测覆盖范围、栖息地面积及个体重叠率等因素推算,相关研究与保护实践普遍认为,当前华北豹的总体规模可能已达到约 600–700 只。
这一估算明显高于 2010 年代中期基于多源资料得出的 174–348 只区间,反映出近十余年来在重点区域内保护成效的累积。

然而需要强调的是,该数值仍主要建立在核心省份(目前主要为山西)的长期监测与模型推算基础之上,全国尺度的系统性普查尚未完成,种群在空间上的高度碎片化和区域差异仍然是制约其长期恢复的关键因素。
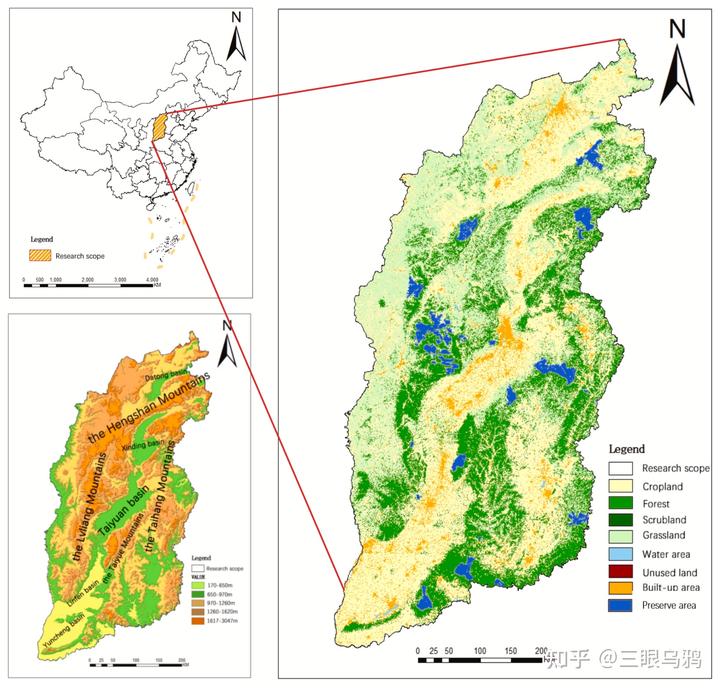

把视角从山西省内监测拉到区域景观尺度,问题会立刻变得更尖锐。
华北地区当前对豹适宜的栖息地总面积仅约 8679km²,且呈高度破碎化格局,被识别为 14 个相对独立的核心斑块,其中仅约 25%被自然保护地覆盖。
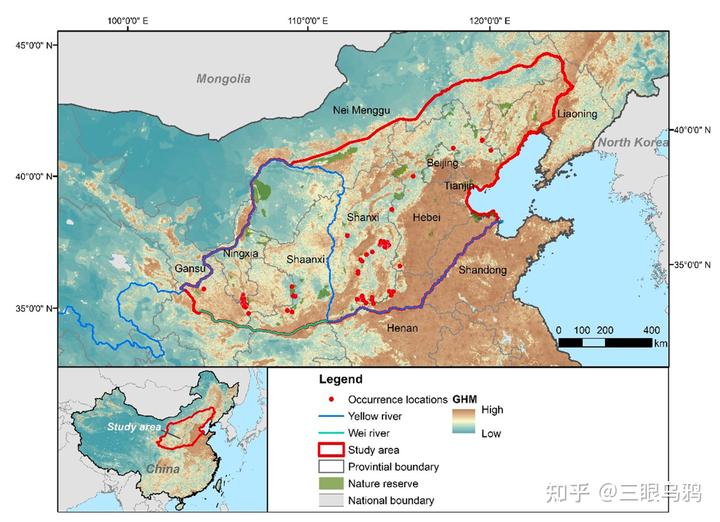
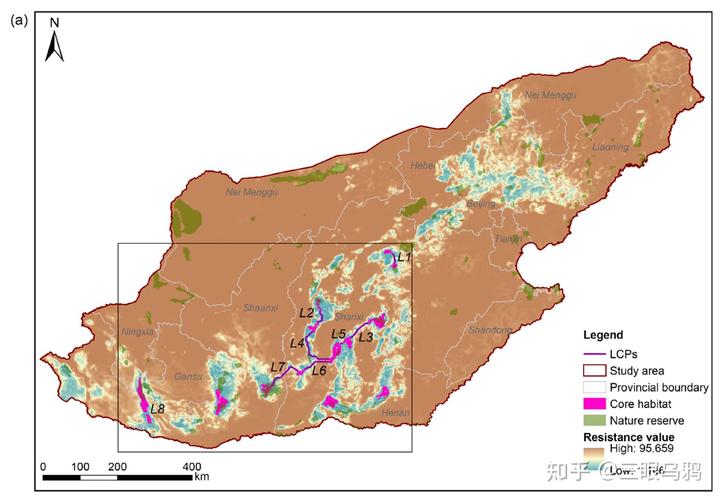
更关键的是,核心斑块之间的连通主要依赖少数脆弱廊道,这些廊道往往处在道路密度较高、人类干扰强烈的地带。
同时,我们要考虑到豹的栖息地偏好:它更倾向于中高海拔、年降水量较高、具有较好森林覆盖的山地环境,并对道路密度和人类扰动表现出明显回避,这意味着扩散通道若落在强干扰带上,其功能极易退化。

当这些脆弱的交流通道被压缩甚至断裂,最先发生的不是个体数量骤降,而是基因交流的枯竭。

2025 年发表的一项群体遗传学研究,利用粪便 DNA 个体识别与群体参数估计,量化了这种看不见的隔绝。

在 2020 至 2023 年间,研究团队在黄土高原东缘(以山西铁桥山为核心)收集了 158 份粪便样本,最终成功从 129 份确认样本中识别出 41 只具体的华北豹个体。然而,基因层面的体检报告却令人担忧。
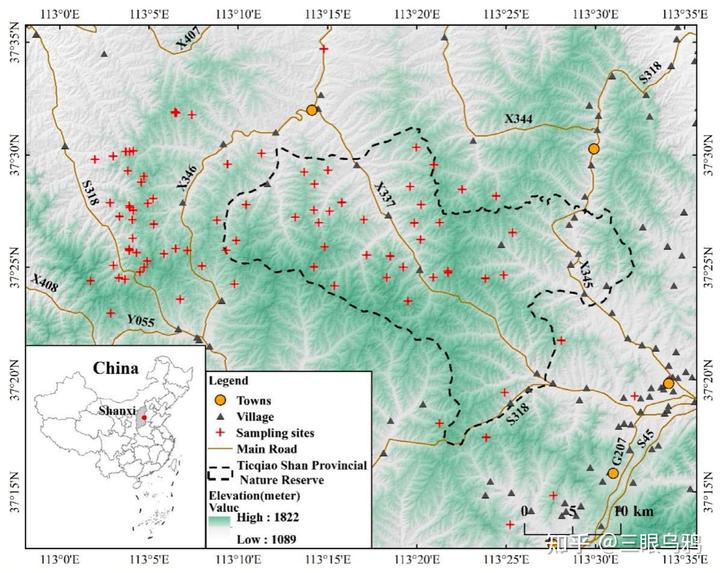
数据显示,尽管我们在野外能看到 41 只豹子,但这个种群的有效种群规模(Ne)仅为 12.20*。简短地说,这相当于维持整个种群基因传递的有效火种仅剩约 12 只,实际参与贡献基因的个体比例(Ne/Nc)只有 0.30。更极端的证据来自母系遗传:约 88%的个体竟然共享同一个母系祖先(线粒体单倍型),这就像一个村子里绝大多数人都来自同一个老祖母的家族,遗传多样性极低。
* 根据保护遗传学的一般准则,Ne 低于 50 通常被视为短期近交风险的警戒线,低于 500 则难以维持长期适应潜力。12.20 这一数值已远低于最低警戒阈值。
这种家族化的直接后果就是近亲繁殖。在识别出的 41 只豹子中,有 32 只都检测出了近交迹象,平均近交水平达到 0.21。虽然计算机模拟(PVA)显示,它们在未来 100 年内暂时没有直接灭绝的风险。
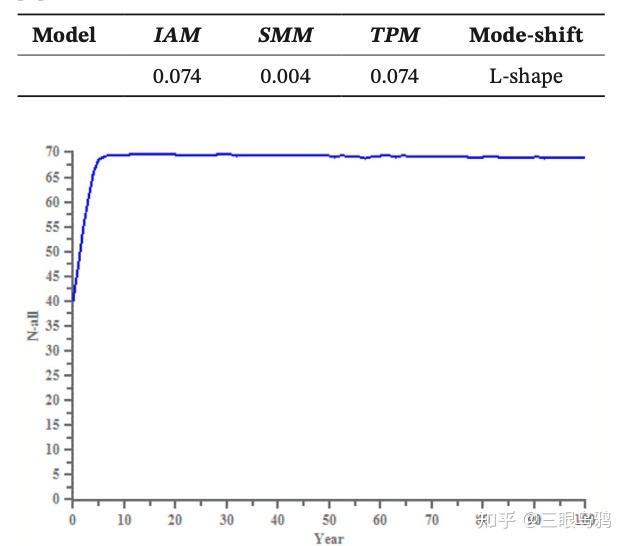
但这更像是一种慢性病:如果缺乏新鲜血液(基因交流),预计百年内它们的遗传多样性将流失约 22%。这意味着,即便豹子的数量在短期内变多了,但整个家族的 DNA 却在变弱,长期适应环境的能力正面临严峻挑战。

正是在这一背景下,北京周边再次记录到华北豹的价值才显得尤为珍贵。它不仅仅意味着北方多了一只豹,更意味着某些曾经断裂的遗传链条正在重新接上。

阜平这种处在核心区与京畿山地之间的节点,一旦被持续利用,就可能成为恢复基因交流的跳板。

华北豹的恢复过程与近年来持续推进的保护措施密切相关,包括加强巡护执法、清理非法猎具、建立红外相机监测网络,以及引入无人机等技术提高巡护覆盖效率。

同时,生境修复也开始从单纯增绿转向结构优化,例如恢复以栎树为代表的乡土阔叶林,为狍子等有蹄类提供稳定食物来源。

山西和顺县的数据提供了有力的佐证:2011–2020 年间,狍子的相机捕获率翻了四倍,与同期华北豹活动频率的上升高度同步,清晰地勾勒出食物网恢复的底层逻辑。
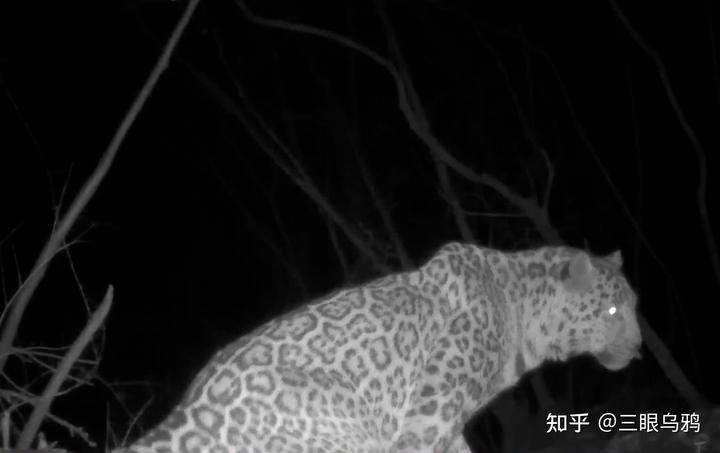
然而,阜平的华北豹影像既是一则好消息,也是一道考题。
华北豹能否真正“回家”,取决于未来十年京冀晋三地能否把当前的保护势头转化为稳定、跨区域的制度安排:包括廊道的长期保护、猎物的系统恢复,以及持续的监测投入。

同时,如何处理潜在的人兽冲突,提升社会容忍度,也是必须面对的现实挑战。

这不仅关乎华北豹这一物种的命运,更是对中国在人口密集区进行生态修复能力的一次关键检验。
如果我们在高人口密度、高土地利用强度的华北,能够重新建立起支持顶级食肉动物生存的完整食物网和生态廊道,那么这一经验将为未来更高难度的物种恢复(如华南虎的重建)提供宝贵的范本。

华北豹的恢复进程,实际上正是为中国未来大型猛兽回归自然,提前进行的一场现实尺度的压力测试。

引用参考
Zhang, M., Wang, C.-H., Zheng, Y.-X., Jiangzuo, Q.-G., Hou, Y.-M., Cao, P., Dai, Q.-Y., Yang, R.-W., Liu, F., Feng, X.-T., Mo, L.-H., & Fu, Q.-M. (2024). Ancient DNA unravels species identification from Laosicheng site, Hunan Province, China, and provides insights into maternal genetic history of East Asian leopards. Zoological Research, 45(1), 226–229. https://doi.org/10.24272/j.issn.2095-8137.2023.292
Dou, H., Liu, W., Feng, L., & Yang, H. (2025). DNA metabarcoding analysis of the North China and Amur leopards’ feeding habits. Current Zoology, XX, 1–10. https://doi.org/10.1093/cz/zoaf027
Wen, X., & Dou, H. (2025). Population genetic diversity and viability of the North China leopard (Panthera pardus japonensis) in the eastern Loess Plateau, China. Ecology and Evolution, 15, e71150. https://doi.org/10.1002/ece3.71150
Zhang, J., Li, F., Zhang, H., & Sun, X. (2024). Conservation strategies for the North China leopard: Establishing effective ecological corridors in Shanxi Province. Ecological Indicators, 169, 112908. https://doi.org/10.1016/j.ecolind.2024.112908
Wang, Y., Liu, M., Xia, F., Wang, Y., Song, D., Liu, Y., & Li, S. (2024). Big cats persisting in human-dominated landscape: Habitat suitability and connectivity of leopards in central North China. Landscape Ecology, 39, 94. https://doi.org/10.1007/s10980-024-01896-y