

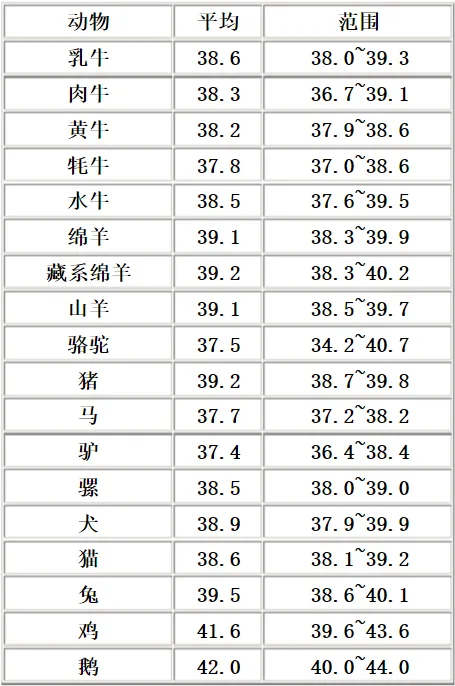
对于人类来说,36~37℃的体温就是最优解,40℃并不一定对我们生存更有利。
探讨体温之前,需要先弄明白一个概念:
进化(演化)是一个淘汰的过程,只有不利于生存的性状(特征)才会被淘汰,凡是不影响物种存亡的特征,都是可能随着基因一代代遗传的。
现在我们再来看看体温的本质:
生物体就是一个巨大的生化反应容器,体温就是这个容器集热所反应的温度。
- 仅仅 1 秒钟,人体内的一个细胞内就会进行 1000 万次耗能的生化反应[1],整个人体高达 6 万亿亿次 /s。
这些消耗的能量,只有少部分转化成了对外的机械功(肌肉效率 10~30%),绝大多数都以热量的形式释放了出去。
这些热量主要以两种方式释放到环境中去。
由于不是本篇重点,仅简单说说
辐射散热:任何有温度的物体都会对外辐射电磁波。
根据斯特藩 - 玻尔兹曼定律:,易知辐射热量与环境温度的四次方差成正比。
对流散热:物体表面流体的相对运动,会带走更多热量。
根据牛顿冷却定律:,易知对流散热与环境温差成正比。
无论散热方式有多复杂,最重要的一点就是:
一个活着的生物,体内的生化反应必须处于不断循环进行的状态,它才不会死亡。
只要存在新陈代谢就会产热,那么它的体温就会比环境更高。
对于低等动物来说,它们具有较低的新城代谢率,例如爬行动物新陈代谢,只有同体型鸟类和哺乳动物的 1/10。
我们可以来简单看看,同体型哺乳动物和爬行动物体温的关系。
如果这个哺乳动物的恒定体温是 38℃(311.15K),环境温度是 20℃(293.15K),当处在无风环境,对流散热足够低,我们可以认为主要散热为辐射散热。
由于它们的新陈代谢率相差 10 倍,且体温与环境温度四次方差成正比。
那么我们可以建立这样的关系式:
哺乳动物和环境温度都是已知的,易得
环境温度是 20℃,也就是说爬行动物的体温,仅仅只比环境温度略高。
也就是说,体温其实是被机体总新陈代谢率所限定的。由于它们的新陈代谢产热,远远环境的热量流通,所以它们也根本没有任何办法,进化出体温调控的能力。
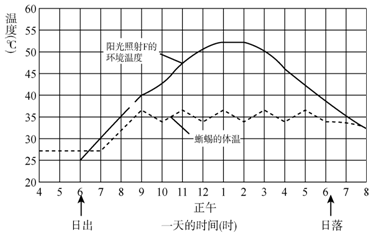
要进化出体温调控的能力,新陈代谢率的提升是最基础的刚需,这是由物理(热力学)所限定。
恒温动物的起源时间,不同的研究有不同的看法。
较晚的一般认为在 1.45 亿~6600 万年前,较早的认为在 3 亿~2.5 亿年前。但无论哪一种观点,都说明恒温的发展是一个渐进的过程。
针对不同物种的研究也表明,不同物种的体温,其实存在多种形式,而不是单纯恒温和冷血之分。例如,有些蝙蝠是异温动物,白天和晚上的体温并不相同,冬眠的动物,存在特殊的体温调节机制,酶和蛋白质都具有适应性的变化[2]。
即便是恒温动物,体温其实也非绝对固定。不仅不同的环境,不同的时间,不同的人体部位体温有所差异,例如,大脑平均温度 38.5℃,核心部位可达 40℃,而骨骼肌在运动时则可以达到 41℃(从这个角度来说,连题主问题的描述,其实也存在一定的问题)。
甚至就连不同生理情况,怀孕、哺乳、运动、发育等等[3][4]不同状态都会对体温造成影响。
其实所谓恒温动物,也仅仅是体温调控能力变得很强,但也并非完全体温恒定。
我们把视角回到演化史来看,物种新陈代谢能力的提升是一个渐进的过程,我们不难推测,在新陈代谢足够低的时候,它们的体温必然受到环境更多的影响,当新陈代谢成倍提升,体温比环境温度更高时,它们便拥有了更多调控体温的可能。
开篇我们就提到,物种的进化是一个淘汰的过程。
为什么新陈代谢足够高的动物,都进化成了恒温动物?
为什么恒温动物中,有的体温更高有的更低,是什么决定了它们的体温?
这可以从两方面的角度来说。
1、什么决定了体温的上限?
认为恒温起源于 3 亿年前的观点,比较核心的支撑,是动物体型大型化与体温的关系。
前文我们提到,爬行动物新陈代谢,只有同体型鸟类和哺乳动物的 1/10。但其实,如果对比不同体型的动物,大型哺乳动物单位体积的新陈代谢,可以比爬行动物更低(当然,总新陈代谢可能高得多)。而大型哺乳动物单位体积的新陈代谢,更是远远比小型哺乳动物或者小鸟低得多,例如大象单位体积新陈代谢仅仅只有老鼠的 1/10,更是只有蜂鸟的 1/100。
之所以存在这样的关系,是因为总新陈代谢与体积(体重)的增加成正比,而散热仄与表面积成反比。
理论上来说,当一个动物高度从 10cm,增加到 1m 时,它的体积增加 1000 倍,新陈代谢也增加了 1000 倍。而它的散热能力,则是增加了 100 倍。它要继续保持散热平衡,那么它的总新陈代谢也只能增加 100 倍。这个总新陈代谢增加的倍数,则是体积增加倍数的 2/3 次幂。
- 这个关系的根本源于几何关系:面积是长度的平方,体积是长度的立方。这其实就是经典的平方 - 立方定律。在工程学和生物工程学上,具有更广泛的运用场景。
也即,某种动物的体积增加 N 倍后,它的陈新代谢率会提升
。
但具体的实验数据,却有所出入。
1930 年代,生物学家克莱伯意图通过研究动物新陈代谢,寻找 2/3 的平方 - 立方规律。然而克莱伯经过统计之后却发现,恒温动物符合的竟然是 3/4 规律,而不是 2/3 规律[5]。
也就是说,体积提升 N 倍后,新陈代谢不是提升
,而是
。
至今,克莱伯定律并没有足够权威的解释。但更多的研究表明,小型动物其实是更加符合 2/3 的平方 - 立方定律的,而大型哺乳动物可能受限于年轻、个体差异,导致不同阶段的单位体重新陈代谢发生了变化,从而更加符合 3/4 定律。
其实,无论是 2/3 还是 3/4 都反映出了这样的规律:
对于大型动物来说,哪怕它们单位体积的新陈代谢效率不高,但随着体积的增加,体内积热的增多,它们的体温也就会逐渐比环境温度高出很多。
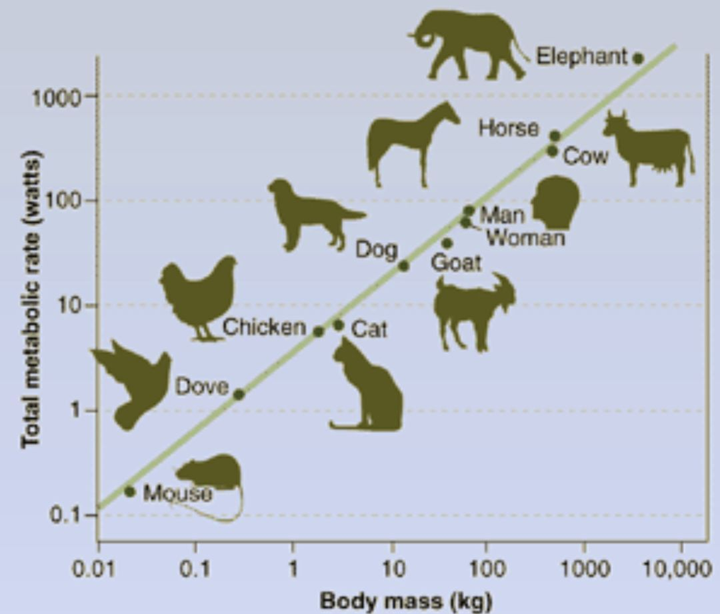
也即,从最基本的热传学角度,我们就能得出:
早期的大型动物(无论大型恐龙还是大型类哺乳动物),体温必然是大大超出于环境温度的。
这也注定了,大型恐龙即便不是恒温动物,也至少是中温动物(介于恒温和冷血之间)。
一般来说,物种的生长率和体温是否恒定具有很大的关系,一般来说冷血动物的生长率足够低,恒温动物的生长率足够高,而大型恐龙的生长率正好介于哺乳动物和爬行动物之间。
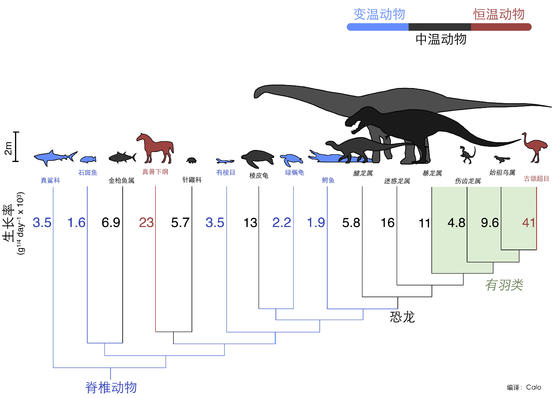
陆生动物大型化,时间恰恰是在 3 亿年前开始。
或许它们一开始,并没有进化出直接调控体温的能力。但很明显体型变大之后,随着积热增加,如果不能调控体温,就意味着环境温度过高时,更容易热死,环境温度过低时,更容易被冻死。
开篇我们就提到,生物体就是一个大型的生化反应器,而催化亿万生化反应需要的是:
酶。
而温度对酶活性,具有决定性的影响[6]。
虽然不同的酶的活性范围很大(0~100℃),尤其是一些嗜热菌体内的酶最高活性温度甚至可高达 80/90℃[7]。但是,不同的生物体内的酶都是为了适应自身环境而存在的。
动物体内酶活性的最适宜温度为 37~40℃,平均温度为 38℃。
正是因为这个原因,绝大多数恒温动物的体温,都在 38℃附近。
另外还有一个美妙的数学巧合,38℃正好位于 0~100℃温度尺度黄金分割点附近。

当一个动物长期处在恒定的环境中,它们的体温也相对恒定,那么它们身体内的各种生化反应也会比较稳定,所使用的酶也会相对固定。
有机体都是由化合物构成,所有生化反应都遵循热力学定律,都遵循阿伦尼乌斯方程:
k=Ae-Ea/RT)。k 速率常数,R 摩尔气体常量,T 热力学温度,Ea 表观活化能,A 指前因子。
这就注定了,温度不仅影响生物体内的化学反应速率,还会引起蛋白质的构象转变、脂类的相变、水的结构变化等等[8]。
这些都注定了,物种从宏观的性状还是微观的生化环境,都对外部环境具有特异适应性。
对于早期的大型动物来说,体温的变化,也令它们身体内的酶也发生了变化。当突然遭遇过高温度或者过低温度时,便可能造成关键的酶失活,影响相应的关键生化反应,从而造成死亡。
在不断淘汰的过程中,那些逐渐能调控体温的个体存活了下来,并得到了繁衍生息。
在不断淘汰演化的过程中,最终促使了恒温动物的出现。
那么最终诞生的恒温动物,它们体内的酶,以及生化反应也总是适应它们所处环境。
从另外一个角度来说,恒温动物的酶在相对较窄的温度范围具有最佳效率,那么所需要酶的范围也更窄。冷血动物的酶则在更大的温度范围发挥作用,不仅需要更多种类的酶,过大的温差,也会造成一些酶在大部分时间都是低效率运行。
例如,一些消化酶最佳活性温度在 38℃附近的,在恒温动物体内总是能高效率运营。而冷血动物,往往只有夏季,这些消化酶才能高效率运行。除此之外,冬眠等特殊的习性,也需要更高的酶成本支持。
也正是因为内部环境对恒温的适应,当突然面临失温,恒温动物更容易被冻死,所以寒冷地带的恒温动物进化出了厚厚的毛发或者脂肪。
在寒冷环境,为了弥补大量的热量损失,机体新陈代谢也会提升,从而具有更高的食物需求,这也导致,恒温动物遭遇食物匮乏时,也更容易饿死。所以对应食物匮乏,一些动物进化出了能量囤积或者冬眠的能力。
恒温其实也并不是总有具有优势,但生物进化过程,总是会为了适应环境,增加新的“天赋”去弥补劣势的地方。每个物种演化,总是一个被环境雕塑的过程,没有某一种特征,是因为它们所处的环境,并没有让他们朝着这个方向演化的生境压力。
人类的演化过程,同样如此。
人类之所以不用像鸟类和蝙蝠那样的进化出 41~42℃的体温(鹅甚至能高达 44℃)。主要还是人类的生境所决定的。我们并没有在体温上,达到它们那样极端的地步。
鸟类可以说是一种新陈代谢成本达到极限的动物,它们通过肌肉煽动翅膀飞翔,随时随地都需要大量的热量供应。
但动物的肌肉效率又十分的低,每对外输出多一份动能,就会产生多 3~10 倍的热量。
由于肉越多基础代谢效率越高,即便不运动,这些肌肉也会消耗大量的热量。
而为了摄入足够多的热量(同时减轻体重),鸟类又缩短了消化道,退化了牙齿,牺牲了消化能力,通过大大提升食物摄入速度,来提升机体所需要的能量。
经过一代代的内卷,一些鸟类甚至进化成需要随时随地地吃,需要随时随地地飞行,甚至雨燕连交配都在空中完成。
其实,它们的高体温根本不是为了对抗病毒而形成的,而是在高内卷的生活状态下,体温必须那么高还能维持热量平衡。其实一些小型鸟类,每当刮风下雨,很容易因为失温而死亡。
地球上的气温在整个地质年代发生了天翻地覆的变化,经历过无数的大冰期。其实对于生物的选择压力来说,低温其实是远远高于病毒压力的。面对冰期的低温和食物缺少的威胁,也注定了绝大多数陆生哺乳动物不可能维持在过高的状态。
人类飞跃式进化的 300 万年,正好经历了第四纪冰期,在寒冷和缺衣少食的大环境下,为了降低代谢成本,会让人类进化出更低的恒定体温。
因此,36~37℃体温不仅没有被淘汰,反而成了大多数。
2、什么决定了体温的下限?
决定体温下限的,首先便是动物最适宜的酶活性温度,这在 37~40℃的范围内。人类其实刚好处于下限。但其实如果考虑原始的哺物动物,体温更低也并不是没有可能。
例如,鸭嘴兽体温仅仅只有 32℃。就像大多鸟类拥有高于 40℃的体温一样,温度稍微超出最适宜酶活性温度,在一定进化压力下也可以合理存在的。
人类没有比 36℃更低,除了酶活性外,还存在两方面的原因:
- 高新陈代谢的刚需:
无论人类还是其它高等哺物动物,哺乳、捕猎、大脑进化等活动的需要,都必须维持高新陈代谢。高基础新陈代谢,注定了体温会处在较高的水平。
- 抵御微生物:
足够高的温度,微生物会失活,这也是题主提问的基础逻辑。
但其实从整个生物界的范围来说,对动物拥有巨大威胁的不仅仅有病毒,还有细菌和真菌。
包括人类在内的 36℃体温以上的恒温动物,很少感染真菌。
有足够的证据表明,足够高的体温,能够让哺物动物抵抗真菌的感染。
例如,兔子拥有 40~41℃的体温,当在兔子温度较低的皮肤或者角膜接种新型隐球菌后,这些真菌并不会向温度更高的区域扩散。当使用糖皮质激素抑制兔子免疫力时,它们则会很快感染而死亡[9][10]。而体温只有 32℃的鸭嘴兽,则很容易被毛霉菌所感染,36℃以上的高等哺乳动物则不会[11]。当一些蝙蝠在冬眠时,随着体温的降低,它们也可能因为感染真菌而死亡。
除此之外,无论两栖动物还是其它冷血脊椎动物,都面临真菌的巨大生存威胁。
研究者模拟真菌感染保护的最佳机体温度,得到的结果是 36.7℃[12]。
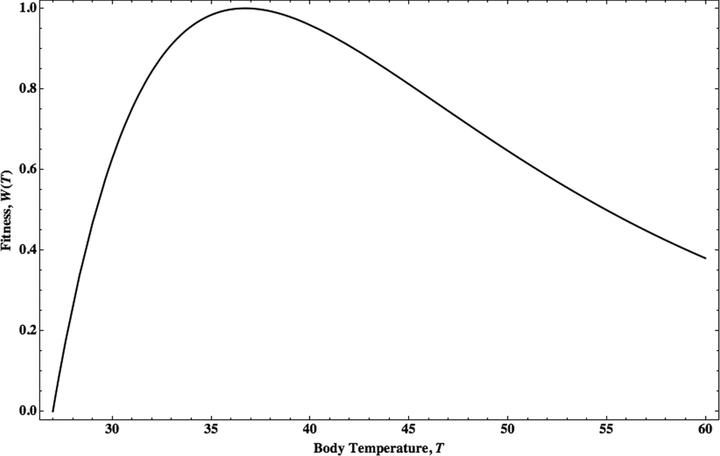
比较巧合的是,这个温度和现今人类的体温比较接近。
说穿了,这个温度是可以足够有效地抵御真菌感染,又具有较低的新陈代谢成本。
统计学表明,人类体温整体降低了,一般认为是现代医学的提升,让人类各类感染风险降低,从而减少新陈代谢成本,降低了体温。
就像怀孕的女性体温会有所升高一样,人类体温本身就和内环境的变化息息相关。
最后要说的是,无论酶活性、基础代谢、体型、摄食、运动,还是免疫需要,人类的体温都是综合的原因决定的。单一的归咎于其中某一项,其实都是不够客观的。
当然,对于某些特殊的物种,可能某些原因会更加的主要。例如,鸟类本身摄食和运动的特化,是造成高体温的主要原因。
由于人类没有鸟类那样的极端,更多的是综合性的影响。