


免费午餐哪里有,南极磷虾在招手。0 耕地、0 饲养、0 添加,只要你抓得到!
第二弹更新,添加延伸阅读:除了鲸鱼,还有谁在吃掉南极的磷虾?
第一弹更新,增加延伸阅读:鲸泵(whale pump)。 顺便解答鲸类种群恢复磷虾够不够吃的疑问。




一年上亿吨的超高再生潜力,蛋白、Ω-3 脂肪酸、虾青素满载,比牛排还营养,比鸡蛋更健康。别人靠喂养、靠化肥,它靠浮游植物自己长。在粮食危机、气候焦虑的今天,磷虾可能就是我们通往未来的蓝色主粮。

南极磷虾(Euphausia superba)是一种生活在南大洋海域的小型甲壳类动物,体长通常为 4–6 厘米,体重 0.5-2 克。尽管个体微小,但其群体数量庞大,构成了地球上最大的动物性生物量之一。根据 Atkinson 等人(2009)和 Siegel(2016)的估算,南极磷虾总生物量大约在 2–5 亿吨之间,其中活跃于水层表面的成体和亚成体就可能超过 1 亿吨。在某些特定水域,例如 Scotia 海、南极半岛西部、Lazarev 海和克尔盖伦高原等地区,其密度高得惊人,有时每立方米海水就可发现数千只磷虾。这种庞大的数量基础构成了它在生态系统和人类食物开发潜力中的核心地位。
南极磷虾的胸肢滤食系统同时具备三种功能:过滤水中颗粒、清理残留物、将食物传送至口器。梳状毛形成密集过滤网;锯齿状末端能刮取并回收前次滤网残留;胸肢泵动配合水流,实现持续高效的进食循环。这使磷虾能在稀薄海水中高效摄取营养,支撑亿吨级种群。
磷虾的胸肢上密布着滤毛(filter setae)与梳状毛(comb setae),共同构成一个精密的“过滤篮”,其网孔大小从 25–40μm 至最细 1–4μm 不等,能有效阻挡较大颗粒并允许水流通过。

磷虾的生命周期大致为 5–7 年,具体取决于栖息地温度和浮游植物资源状况。幼体发育过程包括多个阶段,从卵孵化到成体一般需历时半年以上,期间会经历 10–12 次蜕皮。繁殖季通常发生在南半球夏季(11 月至次年 3 月),雌性可一次产卵数千颗,一个繁殖季中可多次产卵,年总产卵量可达数万粒。在高密度生长区域,年净生物量更新(即种群自然增长)可达 1–2 亿吨(Fach et al., 2002;Tarling et al., 2017),其中部分成为鲸类、鸟类、鱼类等捕食者的食物,剩余部分理论上具有被人类开发利用的可能性。
与人类主要的肉类生产系统相比,磷虾在生态效率和数量上都极具优势。截至 2023 年,全球牛、猪、鸡等家畜总活体重量约为 7 亿吨,其中牛约 15 亿头,猪超 10 亿头。根据 Smil (2011) 和 Bar-On et al. (2018) 的数据,人类的陆地畜牧系统维持这一体量需占用超过 40%的冰冻层外陆地,年耗水量数千亿吨,碳排放占全球温室气体总量的 18%左右。而磷虾则在几乎无人类投入的南大洋中,以浮游植物为食,实现了全球规模的动物性蛋白生产。若将其干重折算为肉类相当营养密度(蛋白含量约为 58–65%),磷虾种群每年可“自然生成”约 0.2–0.4 亿吨的干蛋白产出,约等于全球猪肉年产量的一半,却无需土地、饲料和抗生素等现代畜牧成本。
从农业角度看,主粮单位面积产出远高于自然系统。例如,中国平均稻田产量约为 6 吨 / 公顷,美国玉米带可达 12 吨 / 公顷,单位能量转化效率极高。然而,磷虾分布区域虽单位产量不敌高强度耕作系统,但其覆盖南极环流圈广阔的 1500–2000 万平方公里海域,尤其在特定海域如南极半岛沿岸,其年单位面积可达 10–30 吨湿重(Nicol et al., 2012)。考虑到磷虾无需人为灌溉、施肥与加工,该数字在全球生物资源中依然具有极高性价比。

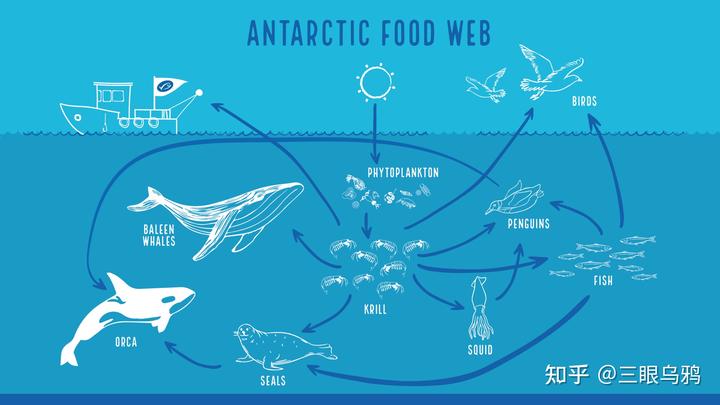
作为生态系统的基石物种,磷虾在维系南极食物网中起到决定性作用。它们以浮游植物为食,并为须鲸、海豹、企鹅、鲱鱼、磷虾鱼等高一级生物提供主要食物来源。例如蓝鲸每日需摄入多达 4 吨磷虾,而一个企鹅群落在一个繁殖季中可消耗数千吨。磷虾的高密度聚集与季节性迁移行为维系了整个南极生态系统的稳定性。研究(Hill et al., 2013)显示,一旦磷虾资源因海冰减少或水温升高而萎缩,整个食物链将发生“崩溃性调整”,从而威胁大洋生态的碳汇功能与渔业结构。
尽管面临重要生态角色,中国已于近年来积极推进南极磷虾的开发与捕捞。根据中国极地研究中心与农业农村部数据,自 2010 年代以来,中国多艘远洋渔船(如“天惠 1 号”“福远渔 868”)已在 CCAMLR(南极海洋生物资源保护委员会)授权下进入南极海域展开磷虾捕捞活动。中国目前年捕量约在 5–7 万吨,位居全球第二,仅次于挪威(约 10 万吨)。但与南极磷虾理论可持续捕捞上限(每年 620 万吨)相比,中国的开发程度仍属初级阶段(CCAMLR 年报,2023)。

对于人类捕捞究竟会不会破坏正常的生态平衡,我们来具体看看。
目前关于南极磷虾资源的担忧,多聚焦于“过度捕捞将破坏生态平衡”,但这一观点忽略了关键事实:磷虾数量波动的主因并非动物捕食或人类活动,而是环境中真正支持其生长的时空极为有限。研究表明,即使磷虾个体总量数以万亿计,其中绝大多数在生命周期早期便因营养不足而夭折。图像数据显示,体长 40 毫米的磷虾,只有在每年 11 月至翌年 3 月的南极夏季才有显著正增长,其余时间受限于低温、弱光与浮游植物匮乏,几乎无法发育或脱壳。
这一“生长窗口”的长度受海冰范围与藻类初级生产力影响极大,并在近年来因气候变化而不断缩短,导致磷虾真正能长成、能繁殖的个体比例远低于理论数量。
下图揭示了南极海域中体长超过 45 毫米的磷虾分布情况。随着体长增加,红点分布范围急剧缩小,>60mm 的个体仅在斯科舍海和南乔治亚岛周边出现,其他大多数海域未曾采集到。这说明,磷虾虽多,但能长成的“成熟个体”高度集中在少数营养热点区域,其种群结构远未充分发挥潜力。

因此,限制磷虾种群数量的并非繁殖能力,而是“营养供给”跟不上“产卵速度”。哪怕产卵量巨大,只要水中缺乏硅藻、甲藻等浮游植物和必要的铁、氮、磷等无机营养盐,99%以上的幼体仍会在数日内死亡。这是一种结构性的生态浪费,而非动态平衡。
从管理角度看,目前全球每年磷虾捕捞量不足总生物量的 0.5%,远未对种群构成威胁。真正值得关注的,是如何突破“短期营养集中—全年饥饿”的结构矛盾,提升磷虾生态转化效率,让这类高效低营养级动物成为更稳定的蛋白来源。

若能在此类区域建设人工干预基地,尝试精确投放无机营养盐或藻类培养物,延长磷虾营养期,如春末或秋初投放硅酸盐、铁离子等,并结合遥感与气候预测手段判断“高产年景”,有望显著提高年净生物量。这类人工干预不仅不会破坏生态链,反而可能缓解鲸类、海鸟等动物的食物危机。

我们不可能一方面向自然索取,又一方面假装不干涉自然,这是掩耳盗铃,也不可能持续。
人类在 20 世纪捕杀了约 290 万头鲸鱼,每年仍猎杀十万以上海豚,早已深刻而永久的改变了海洋生态,如今必须主动干预修复。通过科学手段提升南极磷虾的生态产能,既能满足人类的营养需求,也能为每年万里迁徙至南极捕食繁衍的鲸类海豚等等数量众多的动物提供更充足的食物资源,是责任也是机遇。
中国已经在这个方面积累了丰富的实战经验。
东北虎豹国家公园绥阳分局在冬季大雪封山时,每年定点投放玉米等饲料,保障野猪、狍子等猎物动物生存,为野生东北虎和东北豹提供稳定食物来源。

自 2021 年“十年禁渔”实施以来,中国持续人工投放鲤鱼、鲢鱼、鳙鱼等鱼苗。数据显示,长江干流鱼类单位资源量增长超 16%,监测种类从 200 多种增至 340 余种,生态系统恢复效果显著。
事实证明,袖手旁观行不通,合理规划科学干预才是可持续之道。
磷虾潜力极大、更新迅速,在全球粮食与蛋白短缺日益严重的今天,完全有条件成为蓝色经济的支柱型资源。关键不是“不捕”,而是“科学合理地捕”,同时激活其本就超乎寻常的生态基础生产力。

南极磷虾不仅是自然界中最大规模的动物性蛋白来源之一,还是维系南大洋碳循环和食物链稳定的关键节点。在确保生态平衡前提下,磷虾具备成为人类新型蛋白来源的巨大潜力,尤其可作为陆地畜牧业和传统主粮系统的重要补充。
延伸阅读:
鲸泵(whale pump):从深海归来的肥沃之潮
鲸类在南极捕食磷虾等小型生物,不仅不会减少其总量,反而通过排泄活动滋养海洋,成为重启整个食物链的关键力量。
不少朋友十分担心人类磷虾捕捞对于鲸类鸟类海豹等食物需求的挤占作用,以及未来鲸的种群进一步恢复是否会造成磷虾不够吃的局面,答案是大自然有非常强的调节能力,而且目前南极圈的整个食物链供应主要受限于海水中铁元素等的供应瓶颈而不是捕食压力。
目前南极圈生物总量,远远没有达到上世纪商业化捕鲸前的巅峰。
现代海洋生态学的一项重要发现正悄然改变我们对大型海洋哺乳动物的看法。曾几何时,鲸被认为是“耗费资源”的巨兽,其庞大体型和惊人摄食量令一些渔业利益集团视之为竞争对手。但过去二十年中,尤其是自 2006 年海洋学者 Roman 和 McCarthy 首次正式提出“whale pump”机制以来,越来越多的实地观测和模型研究表明,鲸类实际上在全球海洋养分循环中扮演着至关重要的角色。所谓“鲸泵”,是指鲸类通过垂直迁移和排泄活动将深海富含营养的物质带回表层水体,间接促进浮游植物繁殖,进而带动整个海洋生态系统的初级生产力增长。这一机制已经被视为继“鲸落”之后,人类认知海洋生态系统功能的一大飞跃。
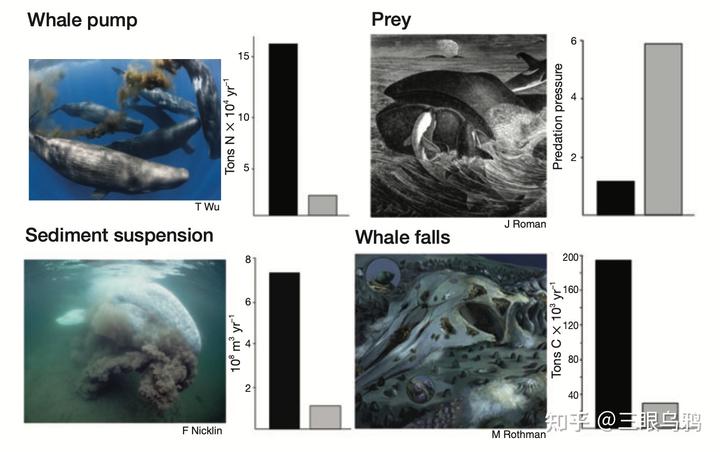
下图以四幅图像和柱状图展示鲸类在生态系统中的关键作用及其因捕鲸造成的衰减。
左上显示鲸泵机制,捕鲸前每年可将 15 万吨氮带回表层,如今不到三分之一;右上显示虎鲸捕食压力因鲸类减少而上升五倍以上;左下灰鲸搅动沉积物曾达 8 亿立方米 / 年,如今作用大减;右下“鲸落”机制原可年封存约 20 万吨碳,现降至四五万吨。整体反映出鲸类所提供的关键生态功能已遭严重削弱。
最早系统描述这一现象的研究是在大西洋北部开展的,研究团队记录到须鲸(如长须鲸 Balaenoptera physalus)和座头鲸 (Megaptera novaeangliae) 在进食后,倾向于游回较浅的海域排泄,释放含氮、磷、铁等养分的粪便。这些排泄物不像一般鱼类迅速沉降,而因富含脂质与微颗粒,在表层形成“营养烟幕”,成为浮游植物繁盛的肥料。在 2006 年的《PLoS ONE》论文中,研究者指出鲸类是“垂直养分回输的重要推动者”,并估算在北大西洋中鲸类粪便可占夏季表层氮输入的 15%以上。
鲸泵机制目前已被观察到发生在多个区域,参与的鲸类涵盖所有大型须鲸种群,包括蓝鲸 (Balaenoptera musculus)、南露脊鲸 (Eubalaena australis)、灰鲸 (Eschrichtius robustus),以及部分齿鲸如抹香鲸 (Physeter macrocephalus)。后者虽不具备完全相同的排泄行为模式,但其狩猎深海头足类后返回表层,也具备显著的“营养上浮”效果。研究显示,一头抹香鲸每天可排泄超过 50 公斤的粪便,其中铁含量远超表层海水平均水平,对海表浮游植物繁殖尤其有利。
这种机制对极地海域,尤其是南大洋南极磷虾带(Antarctic krill zone),意义尤为重大。众所周知,南极磷虾 (Euphausia superba) 是地球上生物量最大的单一物种之一,估算总生物量高达 5 亿吨(随季节浮动),是蓝鲸、须鲸等多种大型鲸类的重要食物来源。然而,这些磷虾的生长与数量波动极为依赖海表浮游植物的繁殖状况,而后者则高度受限于铁等微量元素的供应。由于南大洋本身为 HNLC 区(High Nutrient, Low Chlorophyll),其富含氮和磷,却因铁严重匮乏,浮游植物增长受限,整个食物网效率大打折扣。而鲸泵机制,正是这个系统中铁再分配的重要补充器。
为了更直观理解鲸泵机制的动态过程,研究者对其潜水与排泄行为进行了追踪建模:
下图中鲸在深达 150–200 米的海域反复潜入捕食磷虾,每次上浮到表层后短暂停留并排泄富含养分的粪便。图中的黑线描绘了鲸的连续潜水轨迹,红色虾群代表主要猎物,顶部海面出现淡红色水痕则暗示了粪便在水面形成的养分烟幕。

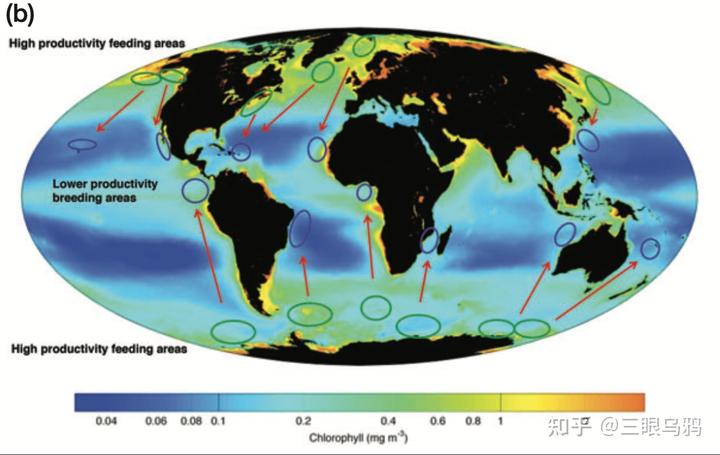
下图展示鲸类迁徙路径与全球浮游植物生产力的空间对应关系。绿色圈标示高生产力的进食区,如南极沿岸、加州与挪威海域;蓝色圈则为低生产力的繁殖区,如中美洲、夏威夷等地。红箭头显示鲸类从高纬度觅食区向低纬度繁殖区迁徙的路线,这一过程不仅连接不同海域,也通过鲸类排泄与死亡行为实现跨洋盆的养分与碳的远距离输送。
以蓝鲸为例,一头成年个体每天摄食超过 3600 公斤磷虾,并排泄大量富含铁的粪便。有研究估算,南大洋在捕鲸前,鲸类种群可维持年均超过 10 万吨铁的回输,而这部分铁可刺激数千万吨的浮游植物生长,间接吸收上亿吨二氧化碳,形成“绿色碳泵”效应。这种天然循环的打断(即工业捕鲸的后果),或许是近年来南大洋磷虾增长受阻、生态系统效率下降的重要原因之一。
与之相对,大众更熟知的“鲸落”(whale fall)机制,指的是鲸类死亡后沉入海底的遗骸成为深海生物的局部生态绿洲。虽然其影响极具象征意义并对深海生物多样性贡献巨大,但从系统层面而言,鲸泵的影响更为持续、广泛,且对表层碳循环有更直接的作用。可以说,“鲸落是死后馈赠,而鲸泵是生者之恩”。
既然铁是限制性元素,那能否通过人为方式补充?科学家曾尝试人工干预以模拟“鲸泵”效应。
2000 年起兴起的人工补铁实验项目,试图通过将铁粉喷洒至 HNLC 海域以促进浮游植物爆发,从而吸收大气二氧化碳、减缓全球变暖。其中最著名者包括 IRONEX、SOFeX、EIFEX、LOHAFEX 与 MESO-Iron 等。

这些项目确实证实了浮游植物对铁输入的高度敏感,例如 2004 年德国团队在 EIFEX 项目中成功制造出一次可持续 3 周以上的浮游植物“人工繁殖场”,浮游植物生物量增长超过 400%。但这些项目也引发诸多争议,例如铁扩散后的生态副作用、藻类死亡后的缺氧带形成风险、生物碳真正沉降深海的效率是否如预测那样高等问题。最终,2010 年《伦敦议定书》* 提出严格管控人工海洋施肥行为,主流科学界开始呼吁回归自然调节路径。
* 正文中提到人工施肥增加磷虾产量和似乎这里矛盾了,其实不然。这一政策主要出于对“人工施肥能否有效增加深层碳汇”的疑虑,并非否定其可能带来的生态层级收益(如促进磷虾生长),两者目标维度不同,正如渔业开发与海洋保护之间需要在协调中寻求平衡。
在这一背景下,恢复鲸类种群的重要性更加凸显。研究表明,南极海域大型鲸类种群目前仅恢复至工业捕鲸前的 10%—15%。若能逐步恢复至历史高峰水平,不仅有助于提升海洋碳汇能力,还有望重启整个磷虾—鲸类—浮游植物之间的良性循环。更进一步,鲸类的迁移路径与气候模式、海流结构交织影响,为全球生态系统提供跨纬度、跨洋盆的养分与基因迁移通道。
当前,有学者提出设立“鲸类养分泵保育指标”(Whale Biogeochemical Services Index),用于量化评估鲸种群恢复对全球生物地球化学循环的贡献。这类指标不仅有助于指导未来的国际保育政策,也提醒我们鲸类并非仅仅是“观赏对象”或“历史遗迹”,而是活跃且功能强大的生态系统工程师。
总之,鲸泵机制让我们重新认识了这些庞然大物的生态价值。在气候变化和生态系统退化日益严重的当下,保护鲸类、促进其自然行为的恢复,不仅是生物多样性的道义要求,也是维系地球生态循环、维持人类自身福祉的必要之举。
鲸类绝不仅仅是巨大的索取者,它和磷虾藻类等构成了共生关系,相辅相成,生死相依。
类似蜜蜂之于花朵,采蜜同时也传播了花粉,促进了植物繁衍。
海洋的可持续未来,正随鲸潮而归来。
除了鲸鱼,还有谁在吃掉南极的磷虾?
南极磷虾虽小,却是整个南极生态系统的能量枢纽;而在鲸鱼数量锐减之后,鱼群、海豹与企鹅迎来了饕餮盛宴,无限量供应下数量暴增。
在地球最南端、被冰雪覆盖的大洋深处,一场历时千万年的生态演化持续上演着。南极圈不仅是一片白色荒原,更是全球最庞大、最高效的寒冷海洋生态网络之一,而其根基,就是一种体长不到 6 厘米、重量不到 2 克的小型浮游甲壳动物——南极磷虾(Euphausia superba)。
磷虾之于南极,就像稻米之于东亚文明,它是整个食物网的核心物质与能量中介,承载着鱼类、鸟类、海豹乃至鲸的生存,是所有高等生物生命链条上的关键环节。


然而这一系统曾经历过巨大的崩塌。20 世纪上半叶,工业化捕鲸活动在南极进入巅峰。根据国际捕鲸委员会统计,1930 至 1960 年代间,南极水域累计捕杀鲸类超过 200 万头。尤其是蓝鲸(Balaenoptera musculus),其南极种群数量一度超过 25 万头,到 1970 年代几近灭绝,数量下降至仅剩 1,000–2,000 头。其他如长须鲸(B. physalus)、南露脊鲸(Eubalaena australis)、座头鲸(Megaptera novaeangliae)等种群也遭到重创。

这一切的直接后果是——巨型顶级捕食者被从生态系统中拔除,食物链顶端产生真空,捕食压力骤降。早在 1982 年国际捕鲸委员会便正式通过商业捕鲸全面禁止令,从 1986 年起南极捕鲸活动大规模退出。此后四十年间,鲸类种群缓慢恢复,至 2020 年代,座头鲸恢复至历史水平的 70–90%,小须鲸超过恢复线,南露脊鲸略有增长,而蓝鲸仍远未回归历史峰值。

Savoca 等人 2021 年研究指出,禁捕之前南极须鲸类年均摄食磷虾量达 4.3 亿吨,而 2020 年代已恢复的鲸类,结合整体估算,2020 年代南极须鲸类每年共计消耗磷虾约 3000-5000 万吨,约为历史峰值 4.3 亿吨的 7-12%。
与此同时,其他生物填补了鲸类留下的生态位空白。南极鱼类、海豹与企鹅等中高层捕食者的相对竞争力上升,尤其在环极流北缘、南乔治亚岛、南设得兰群岛等海域,非鲸类对磷虾的捕食压力持续上升。最新综合估算表明,目前全南极圈各类捕食者对磷虾的年总消耗量约为 1.5-2.5 亿吨。其中,鱼类作为最大消费群体,年消耗量估计在 7000 万至 1.5 亿吨之间;海豹类年消耗约 2800 万吨;鲸类目前年消耗约 3000-5000 万吨(仅为历史峰值 4.3 亿吨的 7-12%);鸟类年消耗约 1000-1500 万吨;而人类捕捞仍处于相对微弱位置,仅 45 万吨左右(2023 年 CCAMLR 数据)。在捕食结构上,鱼类对磷虾的消耗已长期超过鲸类,且因数量庞大、分布广泛、摄食时间延续性强而成为当前食物网中最大的能量流动中介。

鲸类尽管不再是最强摄食者,但依然是最特殊的捕食者。座头鲸目前数量约 30,000 头,活跃于南极半岛西侧(CCAMLR 48.1 区)和斯科舍海,平均每日摄食磷虾 1–1.5 吨,每年集中摄食约 120 天。小须鲸数量超过 50,000 头,体型较小但数量占优,年合计摄食量约 8–9 百万吨。蓝鲸恢复最慢,目前仅 3,000–5,000 头,但个体摄食效率极高,一头蓝鲸每天可摄食磷虾 3–4 吨,主要分布在更深水区域如南大西洋 48.4 区。鲸类捕食主要发生于南极夏季(12 月至次年 3 月),与磷虾生物量高峰期重叠,使其对整个食物链具有“季节性调控”效应。
南极海豹是当前南大洋生态系统中磷虾的重要捕食者,尤其是在鲸类数量锐减之后,其磷虾消耗量显著上升。以食蟹海豹为首的几种海豹,合计每年摄入的磷虾总量可能高达 2700 万至 4000 万吨,超过人类年度捕捞量的数十倍。

其中食蟹海豹的数量估算在 750 万头左右,平均体重约 220 公斤,每日摄食量为体重的 4%,饮食中磷虾比例达到 90%。据此计算,单是食蟹海豹每天便消耗磷虾约 5.94 万吨,年总量接近 2168 万吨,是海豹类中的最大消费者。

南极毛皮海狮约有 400 万头,平均体重 80 公斤,摄食率为 5%,饮食中磷虾占比取 85%,由此得出其每日消耗磷虾约 1.36 万吨,年总量近 500 万吨。

豹海豹种群规模约 30 万头,体重更大(可达 500kg),摄食率 4%,饮食中约 45%为磷虾,其年摄入量约为 75 万吨。
相比之下,威德尔海豹和罗斯海豹等种类虽然数量和体型可观,但其饮食中磷虾比例极低,总体贡献有限。综合上述,整个南极海豹群体每年对磷虾的需求量大致在 2700 至 4000 万吨之间。

这一庞大的摄食规模与 20 世纪中叶鲸类数量骤减密切相关。历史上,鲸类尤其是蓝鲸、长须鲸等大型须鲸,曾主导磷虾的年消耗,单蓝鲸种群就可在鼎盛时期每年消耗数千万吨磷虾。然而,由于工业捕鲸活动,南极须鲸群体整体减少超过 90%,使原本由鲸类占据的生态位空缺下来。食蟹海豹与南极毛皮海狮因此成为最大的获益者,种群扩张显著。食蟹海豹的数量或从几百万激增至当前的千万级规模,成为目前地球上数量最多的中大型哺乳动物之一。这种变化是鲸类种群恢复至今依然缓慢的重要背景,同时也说明了南极生态系统在顶级捕食者缺席下的重构机制,其资源分配已发生深刻改变。
南极鸟类,尤其是企鹅,同样是南大洋生态系统中最主要的磷虾捕食者之一。不同于体型庞大、数量稀少的鲸鱼,企鹅种群在数量上达到千万级。
在所有企鹅中,马克罗尼企鹅、阿德利企鹅和帽带企鹅是三大“磷虾依赖型”代表。

以马克罗尼企鹅为例,全球总种群估计约为 1800 万只,每只体重约 4.2 公斤,日摄食量为体重的 18%左右,其中 85%为磷虾。据此估算,马克罗尼企鹅单日磷虾消耗量约为 11,567 吨,年总量达到 422 万吨。

阿德利企鹅总种群约为 1000 万只,平均体重约 4.5 公斤,摄食率约 18%,磷虾在其食谱中占比高达 90%,其年磷虾摄入量约为 266 万吨。
帽带企鹅在历史上种群估算达到 800 万只,体重略轻,但摄食率相当,饮食中磷虾占比同样达到 90%,年消耗量约为 189 万吨。

这三种企鹅共同构成了企鹅家族中磷虾消耗的主力军,年总消耗合计近 900 万吨。
除了这三种之外,还有巴布亚企鹅和帝企鹅等次要消费者。巴布亚企鹅种群数量较少,约为 77 万只,体重较大(约 6 公斤),但食性灵活,磷虾仅占其食物的三分之一左右,整体年消耗约数十万吨。帝企鹅虽然体型庞大,但种群数量不到 60 万,磷虾只在其食物结构中占次要位置,摄入量较低。

此外,非企鹅类的南极海燕也值得一提,数量可能超过 2000 万只,虽然体重轻,仅 0.65 公斤,但以极高的磷虾比例(约 85%)维持日常能量,其年消耗量也接近 80 万吨。
因此整个南极鸟类系统,特别是企鹅和部分海鸟,每年消耗的磷虾总量稳定在 1000 万至 1500 万吨之间,其中企鹅三巨头贡献了超过 85%。
这些数量与趋势在过去一个世纪中经历了巨大的变化。20 世纪中叶,由于工业捕鲸导致鲸类种群锐减,释放出原本属于鲸的磷虾消费生态位,海豹和企鹅成为直接受益者。尤其是阿德利企鹅和帽带企鹅,在南极半岛、斯科舍海弧等地区的栖息地迅速扩张。通过对企鹅粪便沉积层的研究表明,从 1950 年代到 1980 年代,它们的种群在若干地区增长了一个数量级,部分栖息地由原本的几千对增长至数十万对。此类“磷虾过剩时代”的爆发增长,被认为是企鹅种群历史上最剧烈的扩张期之一。
然而,情况在 21 世纪初期开始出现逆转。一方面,随着气候变化持续推进,海冰迅速减少,尤其是在南极半岛区域,这直接影响了磷虾幼体的生存空间,使磷虾总量呈波动趋势。另一方面,磷虾捕捞业的增长和鲸类逐步恢复也开始对企鹅形成竞争压力。在这一新格局下,原本依赖磷虾的阿德利和帽带企鹅反而成为脆弱群体。据多项长期监测结果显示,这两类企鹅在南极半岛部分区域的数量已下降超过 50%。相比之下,食性更为灵活的巴布亚企鹅反而展现出适应力强的特点,不仅稳定生存,还在向南扩张其繁殖范围,逐步占据原本属于帽带企鹅的冰缘生态位。
综合来看,企鹅在 20 世纪中期通过迅速扩张占据了鲸类留下的磷虾资源空档,是“鲸类空缺期”的最大赢家之一,年磷虾消耗量一度达到千万吨级别。然而,随着生态格局重新变化,气候变暖加速、鲸类回归、人类捕捞增加,这一局面正迅速走向调整期。企鹅的未来生存正面临前所未有的压力,而这一变化也再次凸显出南大洋生态系统的高度敏感性与复杂性。
在南极生态系统中,鱼类是连接底栖生态、浮游动物与高级掠食者之间的关键一环,尤其是数量庞大的中小型鱼类,如灯笼鱼(myctophids)和南极冰鱼(notothenioids),它们在磷虾摄食链中扮演重要角色。根据诸多研究推算,南极鱼类每年消耗的磷虾总量大致在 7000 万至 1.5 亿吨之间,这一数字明显超过了其他掠食者。
最重要的磷虾捕食性鱼类中,排名前三者如下:

首先是南极灯笼鱼(Electrona antarctica),属于世界上最丰富的鱼类群体之一,仅在南极环流带的生物量估计达上亿吨。这些小型鱼每日以体重 10%以上的摄食率持续觅食,主要以磷虾为食,其年磷虾消耗总量估计在 3,500 万至 7,500 万吨之间,占鱼类总消耗量的 50%左右,是鱼类中单一最大消费者。

第二是南极银鲛(Pleuragramma antarcticum),一种广泛分布于大陆架和沿岸的中层鱼类,它是南极鸟类、鲸类和海豹的重要猎物。尽管其总生物量远不及灯笼鱼,但在某些区域的高密度聚集导致其磷虾年消耗量达到 1,500 万至 3,000 万吨,约占鱼类总消耗量的 20%。尤其在罗斯海和威德尔海一带,其生态作用极为关键。

第三是矛尾南极冰鱼(Champsocephalus gunnari),分布在南乔治亚岛和斯科舍海弧地区,是一种中等体型的底层活动性掠食鱼类,对成体磷虾的选择性极高。尽管种群不如前两者庞大,其集中摄食行为导致其磷虾年摄入量达到 500 万至 1,000 万吨,占鱼类总消耗量的 7%左右。
除这三种主要鱼类外,其他南极鱼类(包括其他种类的冰鱼、鳕鱼等)合计消耗磷虾约 1,500 万至 3,500 万吨,占总消耗量的 23%左右。
整体来看,南极鱼类对磷虾的压力主要体现在空间分布广、摄食频率高、繁殖速度快上。尤其灯笼鱼这类具备大规模垂直迁移能力的鱼种,能够在磷虾上层分布带形成高度集中的掠食压力。
三类鱼共同维持着从中层至底层的高效率营养传递系统,使得磷虾能量得以最大化地向上级物种转移。
在这一由磷虾主导的南极圈食物网络中,生态循环的逻辑绝非线性传递,而是一个多层级、多方向的能量回路。从阳光照射至南纬 60 度线以内的海水开始,浮游植物迅速繁殖,尤其在富含铁元素的区域(如冰架融水带与鲸活动密集海域),每年南极夏季(12 月至 2 月)可爆发出大规模浮游植物繁盛带。磷虾作为初级消费者,大量摄取这些浮游植物,将其转化为高密度蛋白和虾青素等营养素。根据 Siegel(2016)数据,南极磷虾生物总量在 3–5 亿吨吨之间波动,个别高年份总生物量可达 6 亿吨,其年再生能力惊人,部分海域的磷虾密度曾在数日内翻倍,维持住整个上层生态系统的能量供给。
磷虾再被鱼类、海豹、企鹅与鲸类广泛摄食,成为营养流向更高营养级的枢纽。值得注意的是,鱼类并非单向摄食者,许多灯笼鱼、南极鱼类同时也是企鹅、海豹与大型鱼类(如巨型鳕鱼Dissostichus mawsoni)的猎物。鸟类与海豹在摄食磷虾的同时,也会竞争鱼类资源,部分季节性或区域性呈现“资源重叠”,形成多物种间的复杂相互作用。这一网络不是固定的金字塔,而是根据季节、水温、海冰、昼夜长度动态调节的流动结构,具备强大的冗余调节机制与局部反馈链。磷虾脱落后形成的“虾雪”沉降至深层,再被底栖鱼类、海参、甲壳类与微生物分解者利用,最终部分营养被再次通过上涌或深潜机制回收至上层。

其中最为神秘而强大的反馈机制之一,来自鲸类。鲸泵(Whale Pump)概念由 Roman & McCarthy(2010)提出,指的是鲸类深潜捕食磷虾后,返回表层排泄的粪便中富含铁、氮、磷等营养物质,可直接刺激表层浮游植物繁殖。在南极圈这样一个铁元素严重贫乏的水域中,鲸泵相当于人为“施肥器”,重启初级生产力。例如蓝鲸每日可排泄数十公斤的悬浮粪便,形成富营养水团带动周边浮游植物爆发,间接增加磷虾的食物供给。Goldbogen 等(2022)测算指出,完整鲸类生态系统的恢复将为南极圈带来年均 13 万吨铁与上千万吨氮的“自然播撒”,远高于当前 CCAMLR 等小规模“人工补铁实验”的投放效率。

另一关键生态机制则是鲸落(Whale Fall)。当一头鲸死亡,其尸体沉入深海后成为深海生物极罕见的高营养输入点。鲸尸上附着的脂肪、蛋白、骨髓可以维持局部微生物、蠕虫、甲壳类、真菌等为期 10–50 年的繁荣生态群落。这些深层生态单位被称为“时间胶囊式生物圈”,是深海极端环境中为数不多的营养稳定源头,同时也将生物圈内的碳、磷、硫等缓慢释放回水体并上升至表层,形成微弱却长期的反馈机制。
正是这两大机制使得鲸类超越了“超级捕食者”的范畴,而成为整个南极圈能量流动系统中的“反馈器官”和“调节因子”。它们通过吃、排泄、死亡三个生物学过程,将深海、中层与表层生态系统贯通,并强化了整个生态网络的“自调节性”和“恢复力”。从历史高峰(蓝鲸 25 万头,年摄食 4 亿吨磷虾)到今天的初步恢复(蓝鲸不足 5,000 头),我们可以清晰看到生态韧性的断裂与重建过程。正如 Savoca(2021)所指出,如果没有鲸类的回归,南极磷虾的生产效率终将受限于铁与氮的瓶颈,进而拖累整个上层生态系统的产出与复杂性。
人类在这个系统中的角色依旧微弱。2023 年 CCAMLR 统计数据显示,年捕捞量为 45 万吨,其中 90%以上集中于乔治王岛、南设得兰群岛等高密度区。
回望整个南极圈生态系统,从浮游植物到磷虾,从灯笼鱼到帽带企鹅,从食蟹海豹到座头鲸,每一个生命节点都紧密相连,构成了一个由能量、营养、行为、空间、时间共同编织的巨大生态网络。
我们必须超越“捕食者 - 猎物”二元逻辑,理解这是一个充满反馈、冗余、交互的多元生态网。通过科学合理的研究与规划,达到人与动物和谐共存的目标。
引用文献正文部分
Klevjer, T. A., Kaartvedt, S., & Aksnes, D. L. (2010). Large scale patterns in vertical distribution and behaviour of krill across the Southern Ocean. Marine Ecology Progress Series, 399, 103–116. https://doi.org/10.3354/meps08315
Atkinson, A., Siegel, V., Pakhomov, E. A., Rothery, P., Loeb, V., Ross, R. M., Quetin, L. B., Schmidt, K., Fretwell, P., Murphy, E. J., Tarling, G. A., & Fleming, A. H. (2009). Oceanic circumpolar habitats of Antarctic krill. Marine Ecology Progress Series, 362, 1–23. https://doi.org/10.3354/meps07498
Siegel, V. (2016). Biology and ecology of Antarctic krill. Springer Polar Sciences. Cham: Springer. https://doi.org/10.1007/978-3-319-29279-3
Fach, B. A., Hofmann, E. E., & Murphy, E. J. (2002). Modeling studies of Antarctic krill Euphausia superba survival during transport across the Scotia Sea. Marine Ecology Progress Series, 231, 187–203. https://doi.org/10.3354/meps231187
Tarling, G. A., Fielding, S., & Klevjer, T. A. (2017). Synchronized swimming behaviour in Antarctic krill: A population-level response to hydrography. Limnology and Oceanography, 62(2), 519–533. https://doi.org/10.1002/lno.10441
Nicol, S., Foster, J., & Kawaguchi, S. (2012). The fishery for Antarctic krill – recent developments. Fish and Fisheries, 13(1), 30–40. https://doi.org/10.1111/j.1467-2979.2011.00406.x
Commission for the Conservation of Antarctic Marine Living Resources (CCAMLR). (2023). Statistical Bulletin – Volume 35 (1993–2022): Krill fishery data. Hobart, Australia: CCAMLR. https://www.ccamlr.org/en/statistical-bulletin
延伸阅读
鲸泵
Blain, S., Quéguiner, B., Armand, L., Belviso, S., Bombled, B., Bopp, L., … & Trull, T. W. (2007). Effect of natural iron fertilization on carbon sequestration in the Southern Ocean. Nature, 446(7139), 1070–1074. https://doi.org/10.1038/nature05700
Bowie, A. R., Trull, T. W., Sallée, J. B., Sallée, J. B., Boyd, P. W., & Blain, S. (2009). Biogeochemical iron budgets of the Southern Ocean south of Australia: Decoupling of iron and nutrient cycles in the Subantarctic Zone by the summertime supply. Global Biogeochemical Cycles, 23(4), GB4034. https://doi.org/10.1029/2009GB003500
Martin, P., van der Loeff, M. M. R., Cassar, N., Vandromme, P., d’Ovidio, F., Stemmann, L., … & Wolf-Gladrow, D. (2013). Iron fertilization enhanced net community production but not downward particle flux during the Southern Ocean iron fertilization experiment LOHAFEX. Global Biogeochemical Cycles, 27(4), 871–881. https://doi.org/10.1002/gbc.20077