
为什么把有蹄类下面的动物分成偶蹄目和奇蹄目而不是 1、2、3 蹄目?

这个问题事实上可能比题主自己想象的还要复杂一点,不仅仅设计到有蹄类哺乳动物的分类问题,更隐含了如何通过观察来认识生物的演化历史,甚至是不是存在一种普遍适用的分类方法而不是长期以来的依靠专家知识或者主观见解。
偶蹄目 Artiodactyla 和奇蹄目 Perissodactyla 两个名字现在都认为是 Richard Owen 在 1848 年提出的,也就是提出了恐龙 Dinosauria 这个名字的那一位。当然,Owen 的工作也是建立在更早的工作之上,但是追根溯源实在是太困难,这里就到他为止了。


有兴趣的同学可以参考 G. G. Simpson 在 1945 年出版的The principles of classification and a classification of mammals
The principles of classification and a classification of mammals. Bulletin of the AMNH ; v. 85
以及 Malcolm McKenna 和 Susan Bell 在 1997 年出版的Classification of Mammals: Above the Species Level
http://cup.columbia.edu/book/classification-of-mammals/9780231110136
具体分类的细节和依据的形态学特征这里就不详细展开了。但是分类学本身就是在快速变化的学科,例如大概上世纪 90 年代提出的鲸偶蹄目 Cetartiodactyla 在 2021 年又被废弃,重新变回了偶蹄目,参考这一篇论文:
On the Unnecessary and Misleading Taxon “Cetartiodactyla”
有蹄类动物分成奇蹄目和偶蹄目也不是一开始就是这样划分的,在 1848 年之前已经有若干种针对有蹄类动物的分类方法,其中比如 Pachydermata 厚皮类(包括象,犀牛,河马,貘等),这些分类方法很多没有包括完整有蹄类动物,也有很多混入了非有蹄类动物,而且很多也没有使用四肢的解剖结构来作为分类的依据。现在看来奇蹄目 - 偶蹄目的划分带有一定的偶然性,毕竟现实中非常难找到一种完全没有例外的形态学分类标准。
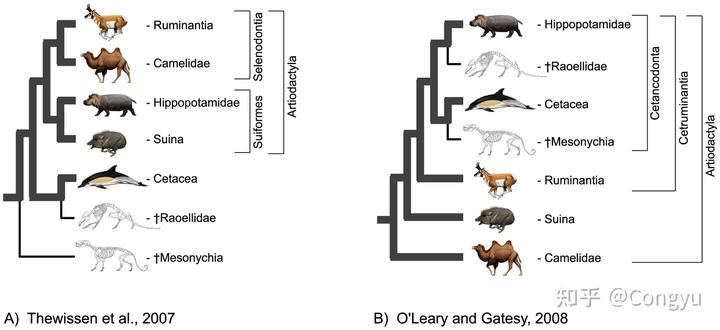
另一方面,随着处于演化历史早期的化石的发现,有蹄类动物里本身就存在一些既不属于奇蹄类也不属于偶蹄类的物种,也就是所谓的干群 stem-group,例如在下面一篇论文中
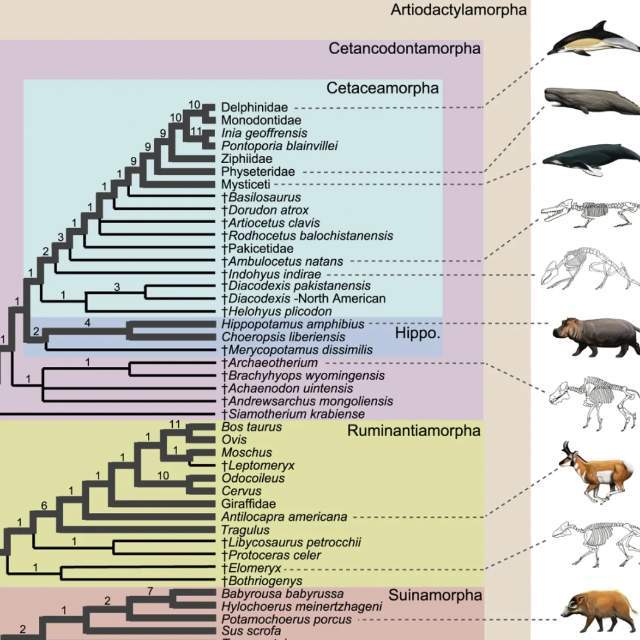
图 1 给出了两种偶蹄目动物演化关系的假说,其中左边的假说中的两个灭绝类群就没有被归入偶蹄目
而图 2 中对更大规模类群的分析,可以看到偶蹄目(严格说偶蹄形态目 Artiodactylamorpha)是上方浅棕色大框,奇蹄目 Perissodactyla 是紧挨着的黄色框,紧接着下面没有底色的几个物种和粉色标记的 Mesonychia 中爪兽类就是有蹄类中既不属于奇蹄类也不属于偶蹄类的物种(“We rooted the trees with the tubulidentate, Orycteropus, as it has been found to be outside of a Carnivora + ungulate clade in a number of studies”)。当然,这样看来有蹄类中的一些早期物种也不一定有蹄。
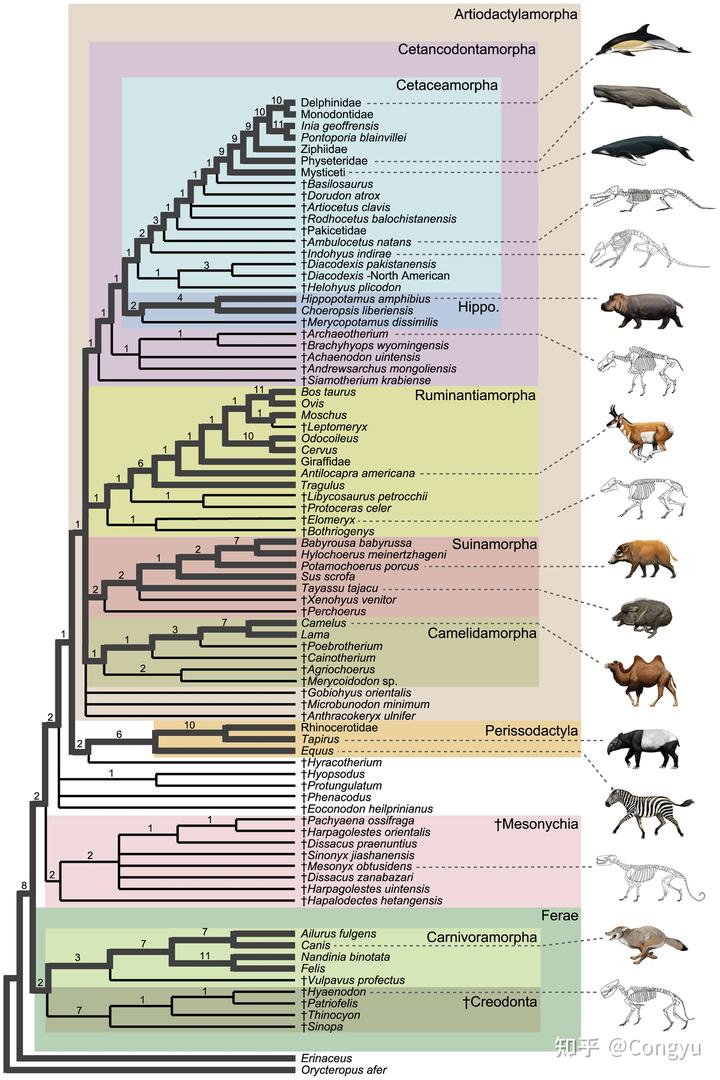
所以小结一下,有蹄类动物只能说现生类群可以分为偶蹄目和奇蹄目,但历史上包含了很多其他的物种,偶蹄和奇蹄作为分类标准存在例外,但是是目前的主流分类方法。1 2 3 蹄的分类早期可能存在,例如 1 蹄的马在很长时间内都被认为独立于其他有蹄类,但现在已经废弃不用。
引申一个问题就是,在(传统)分类中,为什么有大量的基于单一形态特征的二分法存在?例如前面的奇蹄目 - 偶蹄目,后生动物中的两侧对称动物 - 非两侧对称动物,恐龙中的鸟臀类 - 蜥臀类,蝙蝠中的阴翼手亚目 - 阳翼手亚目,被子植物中的单子叶植物 - 双子叶植物。这样的分类方法一方面简单直观,另一方面存在很多例外,而且对于基干类群适用性很差。
首先说为什么不使用三分,四分,甚至更多分的方法,原因之一是任何超过二分的方法都可以等价转换为若干二分法,1 2 3 4 5 蹄就可以转换成 奇蹄(1 蹄 - 非 1 蹄(3 蹄 -5 蹄))- 偶蹄(2 蹄 -4 蹄),多分反而会造成亲缘关系远近的混乱,如果有蹄类分为 1 2 3 4 5 蹄,那么其中 5 个类群的亲缘关系孰近孰远就无法体现。另一个原因是现在对演化历史的假设一般认为物种形成 speciation 就是二分的,实验室里面也许可以人为操作出一些非二分的成种事件,但自然界中应该很难出现,即使有也是极少数,所以实际操作中就忽略掉了。
那么为什么要使用单一特征?在生命演化的历史中,不同特征的演化速率是不同的,比如身体构造是否为两侧对称这个特征在数亿年前就已经被固定下来,尽管存在一些反例(比如发育过程中经历了扭转的比目鱼),绝大多数的两侧对称动物就已经固定了两侧对称的身体构造,不再变动。但如果我们观察一些演化速率很快的特征,例如人体基因组中的单核苷酸多态性 SNP,一个人和父母在同一个基因位点拥有不同的核苷酸,同卵双胞胎似乎也存在不同(没有详细了解过,希望懂的同学评论)?
因此,在对不同尺度的演化历史进行重建的时候,选择恰当演化速率 / 频率的特征是至关重要的。对有蹄类的宏观演化历史而言,是否两侧对称的身体构造或者个体之前的 SNP 都是没有意义的,一个频率太低一个频率太高,相比之下,蹄的数量是一个频率恰当的特征。当然,肯定还存在其他频率接近的特征,比如皮肤结构,头部的装饰角,是否反刍等等都是很好的选择。现在主流观点选择了蹄的数量这个特征,既有各个学者在研究上的努力,也要考虑历史的进程,在上百年的研究和实际应用中被认为是最合适的,尽管现在大家已经看到了这种分类方法的局限性。
最后,目前的分类学依然非常混乱,无论是基于形态还是基于分子的都没有很好的理论基础,还需要更加深入彻底的研究,而不是对现有的系统小修小补,也许和信息论的结合是一个方向。