


目前写了这么多科普以来,大家误会最多的就是“拟态”。甚至有不少人把“拟态”当做神创论的解释。
不如今天就再来好好讲讲拟态吧!
这是一坨鸟屎:

啊不对,其实是台湾铃钩蛾(Macrocilix taiwana),伪装成鸟粪色调的斑点。
但是可以看出这个鸟屎的画法还略微有些幼稚。
我们可以“画”得更好一点:

同为狗屎蛾属,哦不对,是铃钩蛾属的宽铃钩蛾 Macrocilix maia 就更进一步,不但拟态了鸟屎,连鸟屎上面的苍蝇都拟态出来了,嗯,这是一只很有味道的蛾子。
同属的其他蛾子也差不多是这个屎样。
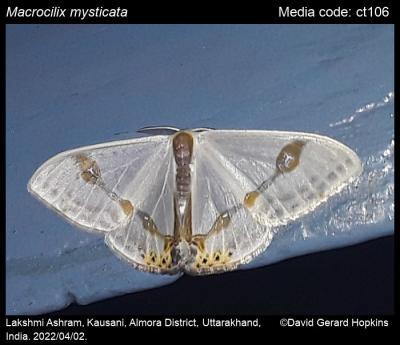
可以看出这个属的蛾子模仿鸟屎还是比较有心得的。
不过这些个屎蛾子只能做出点 2d 的拟态。
柑橘凤蝶幼虫则十分厉害,能拟态 3D 形状的鸟屎:
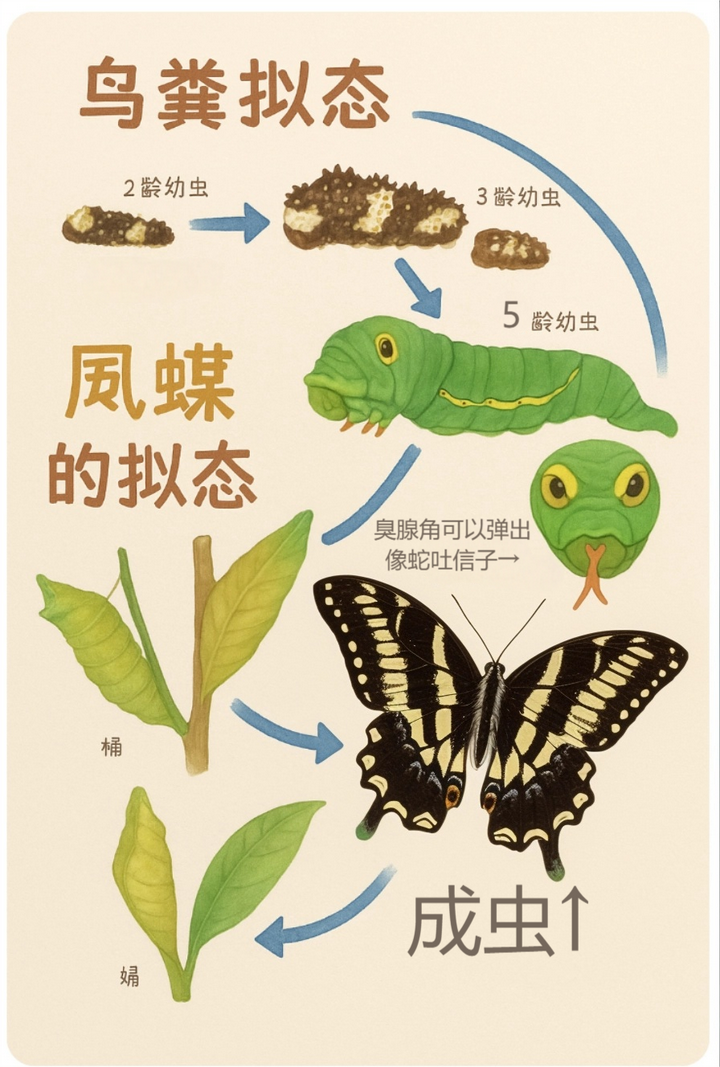
刚开始一龄到四龄幼虫的时候模仿鸟屎是没什么问题的,但是当柑橘凤蝶幼虫长到 5 龄幼虫的时候,问题就来了。
这个时候的幼虫体长有 4cm!而它所处的环境里:
没有鸟!!能拉这么大一!坨!屎!
所以它不得不改变策略,变成绿色,拟态小青蛇:
后胸背两侧有眼状斑纹,腹部有黑色斜纹和白色斑点,并具橙黄色臭腺角(受惊时可伸出散发气味),就像蛇吐信子一样。
当然最后的蝴蝶形态是黑白相间的,跟幼虫时的颜色没有任何关系,毕竟是完全变态昆虫。
另外还有个非常有趣的点,动物拟态植物的时候不仅外貌特别像,而动作也要一致。
模拟植物最常见最出名的应该就是各种螽斯了:

这是亚马逊丛林的螽斯 Acropsis tectiformis,看狐主任手上这一只:

叶脉都拟态得十分清晰。(每只形状略有差异)
还有枯叶系的螽斯:

可以说是达到以假乱真的地步。
但是还没完,刚才说了,光是像还不行,步态也得跟植物一致。
比如我们常见的竹节虫:

你会发现:诶,竹节虫怎么摇摇晃晃的,是不是因为太细了所以下肢力量不稳啊?
其实不是的,大多数拟态成树枝、树叶的动物都有类似的步态,因为这是树叶树枝被风吹动的时候摇晃的姿态,如果用正常昆虫或者其他动物走路的姿势容易露馅。
比如平时是绿色的变色龙(拟态树叶),走路的时候也要模拟这样摇晃的步态:

为什么说这种步态是对树叶的拟态?因为其实变色龙着急早八的时候也是会跑的:

又比如花纹长得像树叶的丘鹬:

bingo,也是一样的步态:

说到这里你可能觉得我越描越黑了,这些精准的拟态怎么看都不像是自然进化的而是被设计的吧?
其实不然。
让我们回到生物课本,看看桦尺蠖的故事:
英国的曼彻斯特地区有一种桦尺蠖,它们白天栖息在树干上,夜间出来活动。该地区原来树木茂盛,树干表面长满浅色的地衣。19 世纪中期以前,桦尺蠖几乎都是浅色的,黑色个体极少。后来曼彻斯特成为工业区,树干表面落满灰色的煤烟灰。到了 20 世纪中期,生物学家们发现,黑色型的桦尺蠖却成了常见种。


从桦尺蠖的颜色变化就可以看出生存压力所产生的自然选择对生物的外观有多大的影响。
拟态是出于这样的压力,由自然选择所决定的进化策略。
拟态分为三种:
- 贝氏拟态(Batesian mimicry,以亨利·沃尔特·贝兹命名),一个物种拟态模仿另一个成功的物种,显得有毒或者是无食用价值。但该拟态并不会攻击捕猎者。
- 穆氏拟态(Müllerian mimicry,以 Fritz Mülle 命名),一个物种以鲜艳的体颜等手段警告捕猎者其毒性或不可食用性,如果捕猎者执意要试试,那么它就会得到教训,例如捕食者从一次失败的捕猎中很快认识到猎物有毒。
- 进攻性拟态(Aggressive mimicry),拟态成为无害的物种以吸引猎物。

贝氏拟态是防御性拟态,这种拟态不会攻击捕食者,只是逃避捕食。
像刚才说的拟态鸟屎和树叶等就属于贝氏拟态。
能够拟态某种其他的前提是:该种动物具有足够多的可以变异的位点。
众所周知,昆虫数量非常之多,大概有 200 多万种。
而昆虫通常还具有极高的繁殖率,一次可产下成百上千甚至更多的后代。
这种“以量取胜”的策略,在演化上具有重要意义,而且基因复制过程并非完美无缺,总会随机出现错误,即基因突变。
庞大的后代数量,如同购买了海量的“基因彩票”。虽然单个突变发生的概率极低,且大多数突变是无效甚至有害的,但巨大的基数使得有益突变的出现成为可能。
这些变异位点(基因座),在生存竞争异常激烈的自然界,任何一个微小的、能够提升存活率或繁殖机会的突变——例如,翅膀花纹偶然更像一片树叶,或体色恰巧与有毒的同类相似——都可能成为这个个体及其后代的“幸运符”。这个幸运的个体更容易躲避天敌或成功捕食,从而有更多机会将这一有利变异遗传给下一代。经过代代累积,这种微小的优势不断被放大,最终演化出令人惊叹的逼真拟态。
另外,除了可变异的基因要足够多以外,环境也会极大的影响动物们对拟态的进化选择:
以 2023 年对竹节虫的研究为例:
加州有三种Timema 属的竹节虫,它们是无翅的植食性昆虫,依靠拟态成树枝树叶的形状隐藏自己。它们通常有绿色与黑色(深色)两种色型,不同种的色型差异程度不同:
- T. cristinae 和 T. bartmani:色型高度差异化;
- T. chumash:色型较为连续。

科学家研究发现植物颜色变化越离散(如强烈的绿与棕对比),竹节虫色型的差异程度就越高。
- 色型差异性顺序为:T. cristinae > T. bartmani > T. chumash
这几种竹节虫基因库的复杂度是类似的,但是根据所处环境的不同,它们基因的变异程度也不同。
控制着虫子的体色的遗传区域叫 Mel-Stripe ,它决定竹节虫是绿色还是深色。
研究者关心这个区域的杂合度,也就是绿色和深色基因是否常常出现在同一个个体里。
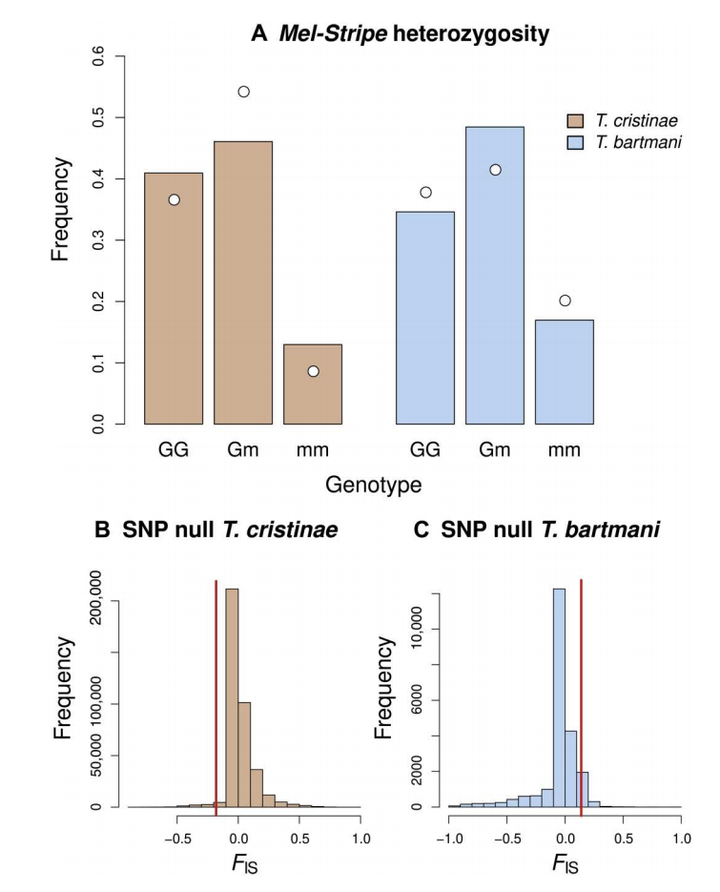
1. 杂合个体是多了还是少了?
- 在 T. bartmani 中,杂合子(即同时有绿色和深色基因的个体)比预期少。这说明绿色和深色型可能不太愿意互相交配,或者交配后杂合子生存率低,这可能是形成物种隔离的一种信号。
- 在 T. cristinae 中,反而是杂合子多了,比预期还多。这可能说明:这种虫子在某些情况下,杂合子生存得更好(异型优势)。
2. 跟全基因组相比,Mel-Stripe 的差异更明显吗?
研究者把 Mel-Stripe 这个区域和虫子整个基因组中的其他几万个基因位点做了比较:
- 对 T. bartmani 来说,Mel-Stripe 的杂合子缺失虽然不是最极端的,但比全基因组大多数区域的情况更严重,说明这个现象有进化学上的意义。
- 对 T. cristinae 来说,Mel-Stripe 的杂合子过剩非常明显,比全基因组其他区域都极端,说明这个区域在杂合子中起着特别的作用。
所以研究者最终的结论是:
在 T. bartmani 中,绿色和深色虫子可能已经有部分隔离,不太愿意“通婚”;而在 T. cristinae 中,绿色和深色基因“合作愉快”,杂合个体甚至可能有优势;
这个 Mel-Stripe 区域可能在某些物种中变成了一个超基因(supergene),帮助维持色型的稳定分化。
希望上面这段阅读愉快,如果没有很懂,那我解释解释:
就是说在自然环境下,这些竹节虫本来就有体色变异的自然倾向,但是在植物对比度更高,颜色更多的情况下,不同颜色的竹节虫更愿意通婚,所以它们后代颜色也更复杂,更接近各种植物颜色。
而植物颜色差异不大的地区,不同颜色的虫子不愿意通婚,因此后代的颜色也比较保守,差异性不大。
这就引出拟态选择的另一方面:

除了本身作为保护色以外,性选择等因素也会加速拟态的筛选,让优势颜色更快的适应环境,以及加速繁衍。
以上~
参考文献:
1morphs in Timema stick insects Disruptive selection and the evolution of discrete color