


不知道大家还记不记得,在学习植物学藻类部分的时候(陆时万主编的那本经典绿皮上下册教材),第一个印象是藻类部分的分类感觉特别“朦胧”,第二个印象就是它们特别奇葩的细胞器,特别是质体,裹了一层又一层的膜结构,像切开的洋葱一样。
然而当年藻类大概是最不受重视的部分,很多学校可能压根儿就不讲吧,所以当时只觉得混乱奇葩,各种无人问津的未解之谜系列,也没有多作仔细深入思考。
谁能想到到现在关于藻类的系统分类研究和细胞研究,已经彻底刷新了我们的三观。藻类早已不是当年那个混沌模糊被一股脑儿塞到一起去的类群了。有些藻类变成了绿色陆生植物的亲戚,有些藻类变成了疟原虫的亲戚,有些藻类变成了原来属于真菌的水霉的亲戚。
目前藻类的分类到底是什么样,可以阅读我的这篇回答:
地球上最初的生命为什么有的进化成了植物有的进化成了动物? - 知乎
也方便大家熟悉一下下面出现的各种闻所未闻的类群名称。(好像那个答案最近被某公众号拿去做成视频了,还挺火的,然而我并不之情 ORZ。。。嘛。。。)。
接下来就要给大家讲讲藻类里面为什么会有那么多重重叠叠的膜结构,也就是内共生和二次内共生学说。
内共生学说认为真核细胞中的质体(包括叶绿体)和线粒体来源是与其共生的原核细胞,后者被前者吞噬并逐渐演化成现在的细胞器。内共生学说的已经有一百多年的历史,发现发展到现在已经是很成熟并被广泛接受的学说了。70 年代 Bonen 和 Doolittle 发现了质体和蓝细菌核酸之间的相似性;80 年代 16S 核糖体 RNA 的同源性分析更加证实了质体的蓝细菌(cyanobacteria)来源(Giovannoni et al., 1988)和线粒体的变形菌(proteobacteria, 包括大肠杆菌,霍乱弧菌)来源(Yang et al., 1985)。
质体和线粒体通过内共生到底起源了多少次?目前认为两者都是单起源的,内共生只分别发生了一次。其最主要的证据来自细胞器外蛋白质输入装置( protein import apparatus )的同源性(Dolezal et al., 2006; Mcfadden et al., 2004)。如果内共生发生了多次,应该会演化出很多套独立的蛋白质输入装置。但所有真核生物的线粒体都共用一套转运装置(TIM, TOM, trans-locon of the inner/outer mitochondrial membrane. Zarsky et al., 2012; Shiflett &Johnson, 2010),质体也共用一套转运装置(TIC, TOC, translocon of the inner/outer chloroplast membrane. Bullmann et al., 2010; Shi & Theg, 2013)。这一同一性支持质体和线粒体的单起源。
蓝细菌通过内共生(primary symbiosis)演化成质体的时间大概在 12 亿年前(Parfrey et al., 2011),其后在某些藻类中发生了二次内共生(secondary symbiosis)。要注意这里并不是说通过内共生产生了新的质体,而是在质体产生后,某些真核细胞通过吞噬已经具有质体的其它真核细胞,获得了结构更为复杂的质体。其主要证据是某些藻类的质体被三层或更多的膜结构(一般为四层)包裹(Gibbs, 1981)。
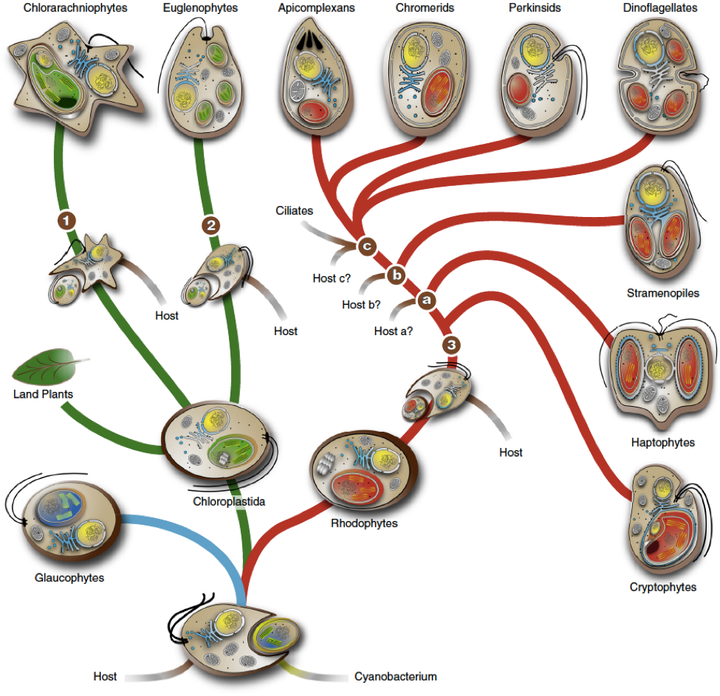
(Zimorski et al., 2014)
这幅图比较清楚的展示了来源于吞噬泛植物(Archaeplastida)细胞的二次内共生演化。泛植物的共同祖先通过吞噬蓝细菌获得了质体,然后演化出三个类群:
1.灰胞藻(Glaucophytes)的质体有肽聚糖层,是蓝藻细胞壁的遗留,也是内共生假说最有利的支持证据之一。
2.绿色植物(Chloroplastida)。包括绿藻和有胚植物。目前发现了两次独立起源的二次内共生是通过吞噬绿色植物细胞进行的。一次发生在古虫界的裸藻(Euglenoids,包括眼虫,就是那种可以进行光合作用的原生生物)中,另一次发生在有孔虫界的网绿藻( Chlorarachniophyte )中。
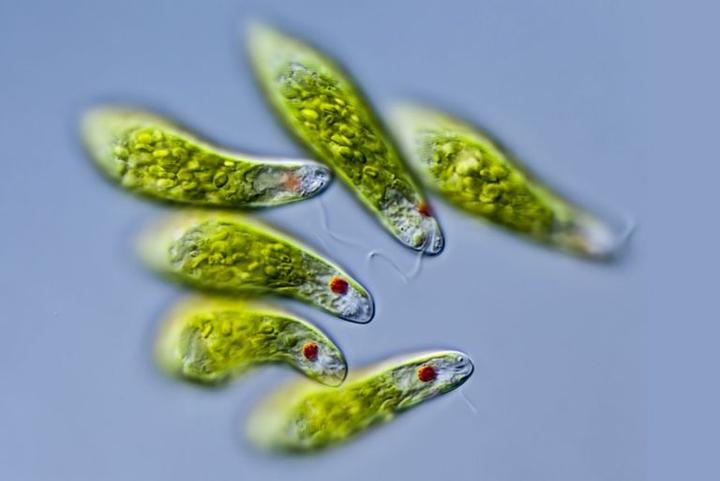
眼虫(Euglena)
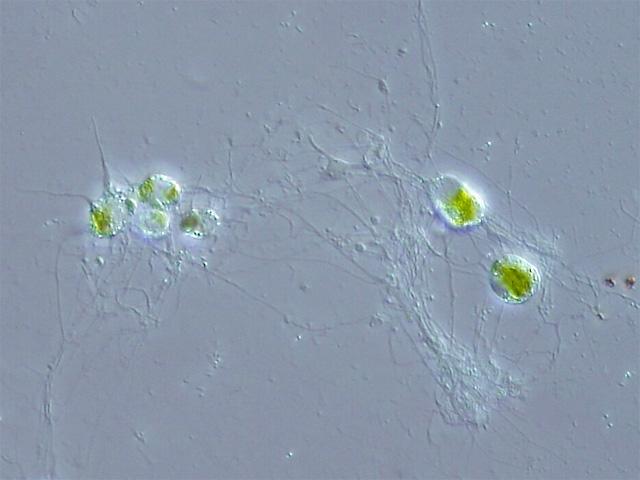
网绿藻(Chlorarachnion reptans)
3.红藻(Rhodophytes)。通过吞噬红藻类细胞进行的二次内共生则要复杂得多。目前已知的包括隐藻(Cryptophytes),定鞭藻(Haptophytes),不等鞭毛类(Stramenopiles,包括金藻,褐藻,黄藻,硅藻,卵菌),囊泡虫类(Alveolates,包括疟原虫,草履虫,甲藻)。现在还研究得很不透彻,不同分子系统学得出的结论争议非常大,到底独立起源了几次也不清楚。Maier 的研究团队通过与论证质体和线粒体单次起源相同的逻辑,考察了位于红藻类质体最外层第四层膜和第三层膜之间的蛋白质运输系统 SELMA(symbiont-specific ERAD(endoplasmic reticulum associated degrdation)-like machinery),并发现它们是同源的(Sommer et al., 2007; Bolte et al.,2009)。所以红藻类二次内共生的藻类来源可能是单一的。但我们仍然不知道到底有多少宿主独立获得了这种红藻细胞。
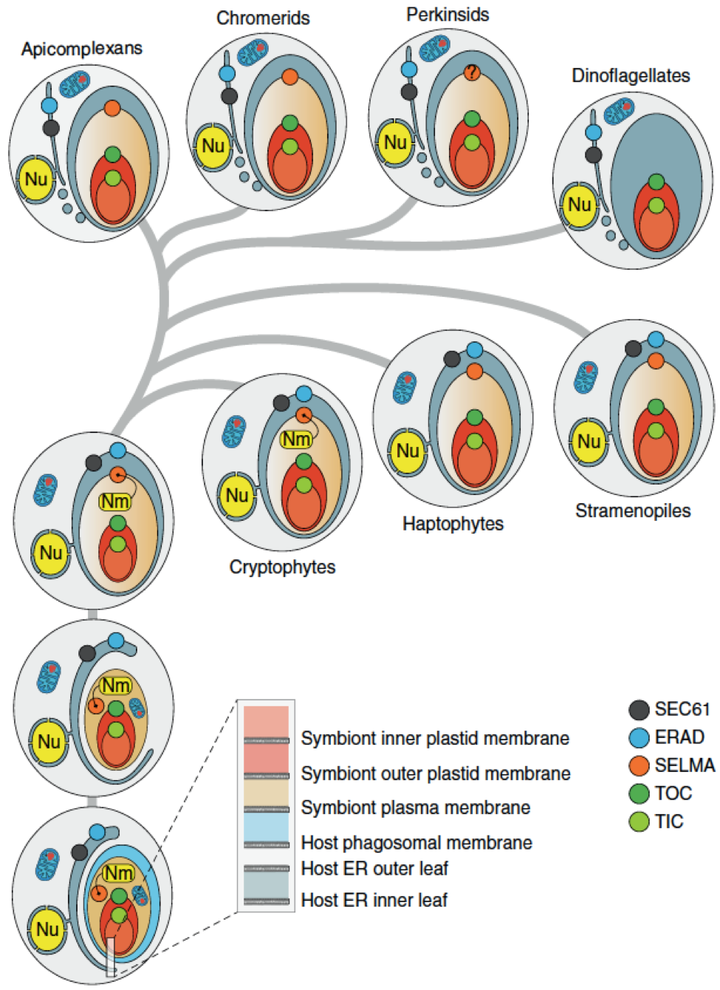
红藻类二次内共生膜和蛋白质运输系统的演化过程(Zimorski et al., 2014)。蛋白质运输系统一共有三个层次:TOC 和 TIC 是红藻细胞内质体的运输系统;SELMA 是红藻细胞自己的运输系统;ERAD 和 SEC61 是二次内共生宿主细胞自己的运输系统。
被吞噬的红藻细胞首先被宿主细胞产生的吞噬体膜包裹(phagosomal membrane)。吞噬体膜消失后被宿主细胞本身的内膜系统包裹。值得注意的是在隐藻(cryptophtes)质体中还存在核型体( nucleomorph,Nm ),是一种位于两层膜结构之间的细胞核残余结构,表达 SELMA 相关蛋白质,应该是被吞噬红藻细胞本身细胞核的遗迹,也进一步佐证了内共生假说。其它各类群这一结构都彻底消失了。
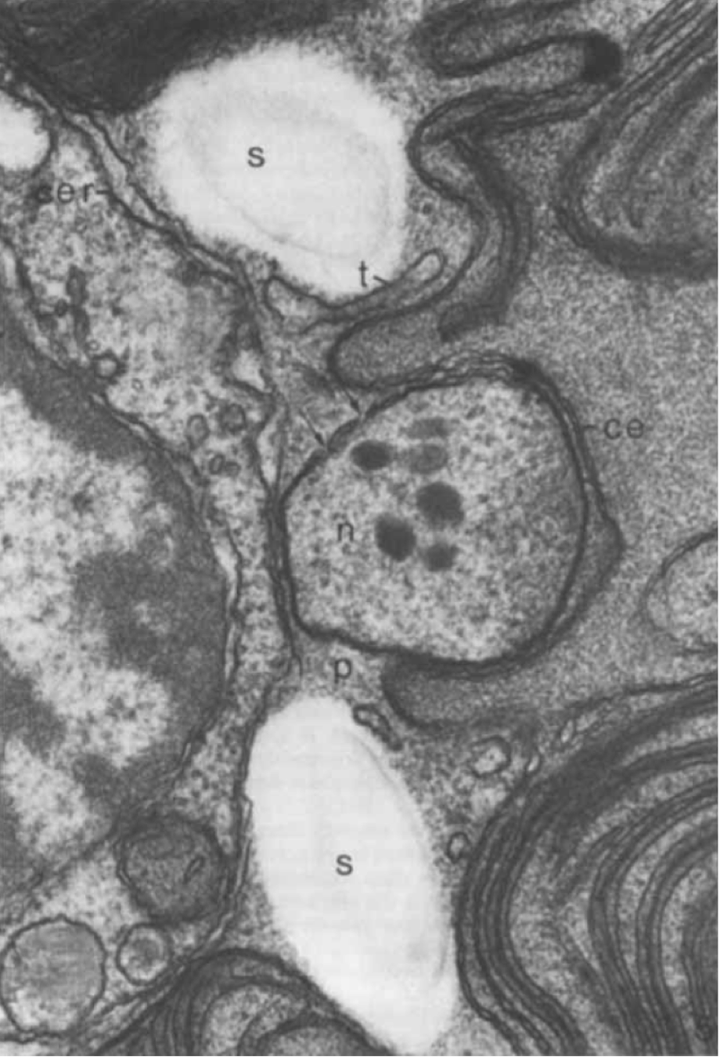
蓝隐藻(Chroomonas salina)的质体超显微结构(Gibbs, 1981)。图中 ce 是质体内部两层膜,cer 是质体外部两层膜,n 是核型体,位于两套双层膜系统之间。核型体本身也被膜结构包裹,箭头所指部分是类似于核孔的部分。
含有多甲藻素的双鞭毛类(Dinoflagellates,包括甲藻)跟其它类群不同,只有三层膜结构,也没有 SELMA 系统。应该是在演化过程中丢失了。
延伸阅读:Zimorski V, Ku C, Martin WF, Gould SB. 2014. Endosymbiotic theory for organelle origins. Current Opinion in Microbiology22:33-48.
几位作者都是常年专注于研究内共生的学者,发过不少 Nature 和 Science。这篇比较新的综述总结了最近这个领域的新进展和还未解决需要继续研究的问题。