



从演化选择压力的角度看,长颈是一条生理成本高、生态位窗口窄的小路。

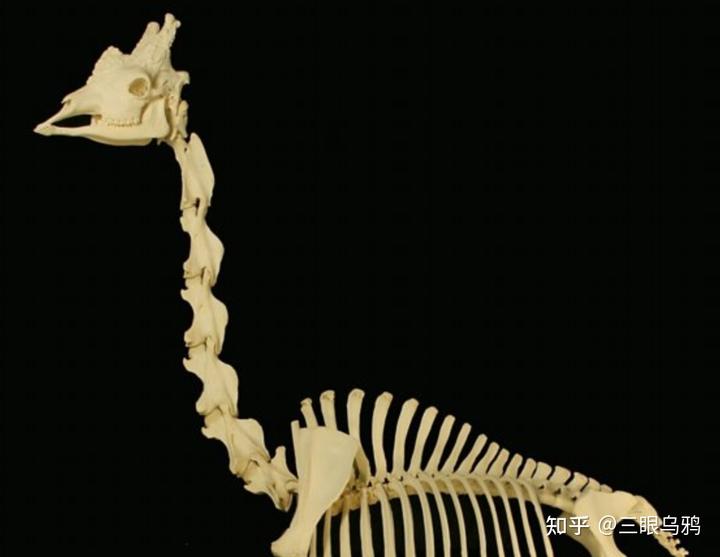
对比长颈鹿、马和牛三条演化路线,会发现决定性因素并不在于能不能拉长脖子,而在于拉长之后是否能在长期竞争获得更大的竞争优势并抵消其昂贵的代价。


马的演化核心始终围绕低位取食与高速奔跑展开。
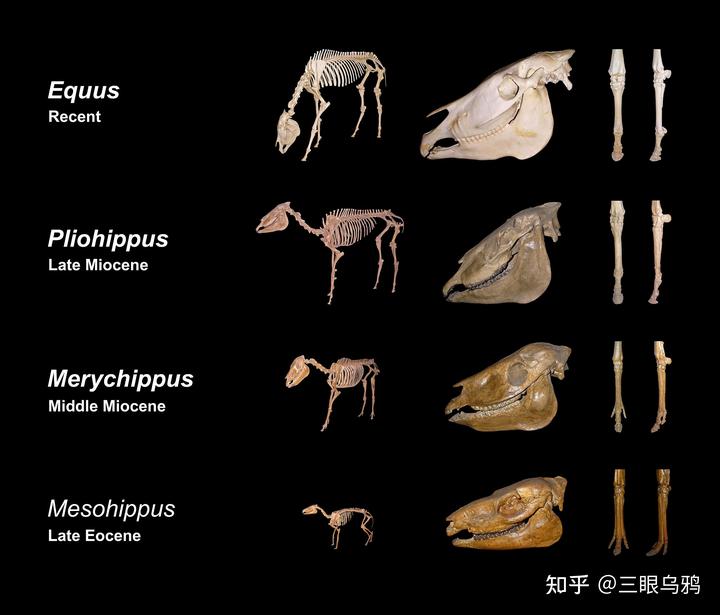

自中新世草原扩张以来,马科以贴近地面的啃食为主。这里存在一个严格的物理约束:马的颈部长度必须与延长的四肢相匹配,才能确保嘴巴能接触到地面。 一旦超过这个够得着地的必要长度,继续加长颈部对马来说就是纯粹的负累。


马对速度、稳定性和能耗极度敏感,颈部作为前躯的重要摆动质量,过长会显著增加转动惯量,削弱高速奔跑和急转能力。因此,马选择了长腿 + 适度长颈的组合,依靠群体警戒和奔跑来解决生存问题,这比演化出去吃高处树叶的长颈要划算得多。

牛的方向看似不同,但结论一致。牛科的优势建立在反刍系统和高效率、长时间的低位取食之上。
牛科动物普遍拥有沉重的头部附属物(角),这构成了巨大的力学限制。 头部本就沉重,若颈部显著拉长,力臂增加,维持姿态的能耗和颈椎压力会呈指数级上升。



此外,牛科内部的竞争主要体现在体型对抗和角的冲撞上,这种格斗模式要求颈部粗壮短促以提供稳固支撑,与纤细灵活的长颈演化方向背道而驰。




长颈鹿之所以成为例外,是因为多重条件罕见地同时成立。
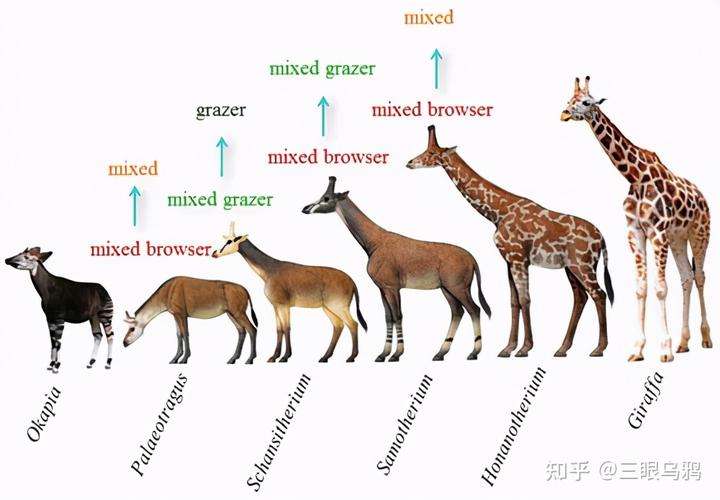

非洲稀树草原中存在稳定的垂直食物分层,高位树叶长期竞争较弱,长颈提供了不可替代的独占资源。
斑马主要取食贴近地面的高纤维禾草,取食高度通常不超过 30–40 厘米,依赖的是数量庞大但质量较低的草本资源。

多数羚羊则处在中间层级,它们既啃食草本,也取食低矮灌木和幼嫩枝叶,取食高度大致集中在 0.5–1.5 米之间,通过灵活性和选择性在草与灌木之间切换。
但由于身高和体型的限制,很难长期进入 2 米以上叶层。


而长颈鹿的取食高度则显著跃升,常规取食范围在 2–4 米,必要时甚至更高,这一高度区间中的树叶,尤其是金合欢等乔木的叶片,长期处于竞争压力极低 * 的状态。
*此处所指为反刍类内部的长期竞。大象主要通过偶发性取食或破坏性方式利用乔木资源,其取食策略与反刍类不同,不构成直接竞争。季节性或极端干旱属于时间尺度更大的环境约束,而非常态条件下的资源竞争。

同时,长颈鹿并不以极限奔跑速度为生存核心,且其角部退化为较轻的骨质突起(ossicone),减轻了头部配重。

而且体高本身即可降低被捕食风险(长颈鹿的踢击非常有力),再加上角的退化,从而为颈部结构延长让出一定的力学与能量空间。

更关键的是,雄性之间的甩脖搏斗(Necking)使颈部结构直接进入了性选择通道。在一定程度上对颈部延长产生了持续的定向强化。

即便如此,长颈鹿的近亲并未普遍走向长颈化,说明只要垂直食物分层、捕食压力结构或性选择机制中的任何一环发生变化,长颈方向都会迅速失去优势而被淘汰。

长颈的稀有并非偶然,而是强约束下的必然结果。马和牛的成功,正是建立在避开这条高成本、高风险路径之上。
而长颈鹿的存在,则是在多重条件罕见重合的前提下,才得以实现的一次高度特殊的演化结果。
而且这个罕见的成功不是没有代价。

首先是生理成本:为将血液泵送到高处头部,长颈鹿必须长期维持极高血压,其心脏结构和能量消耗远高于一般哺乳动物,这是持续一生的负担。

其次是行为与安全代价,长颈鹿低头喝水时必须分开前肢、姿态笨拙,警戒能力显著下降,是捕食风险最高的时刻。

再次是生态限制,高大的体型与长颈使其难以进入密集丛林或灌木环境,在空间复杂、遮蔽性强的栖息地中反而不利。

最后是运动学代价,长颈与长腿组合降低了快速转向与灵活性,使其并不适合高机动逃逸。正因这些多重成本并存,长颈成为一种极端而罕见的演化结果,只在特定环境条件下才得以维持。

在进一步详细讨论不同草食兽颈部形态差异之前,有必要先厘清一个核心问题:
推动长颈化的主要力量究竟是什么:是取食高度带来的生态收益,还是甩颈搏斗等性选择机制的影响更为关键?
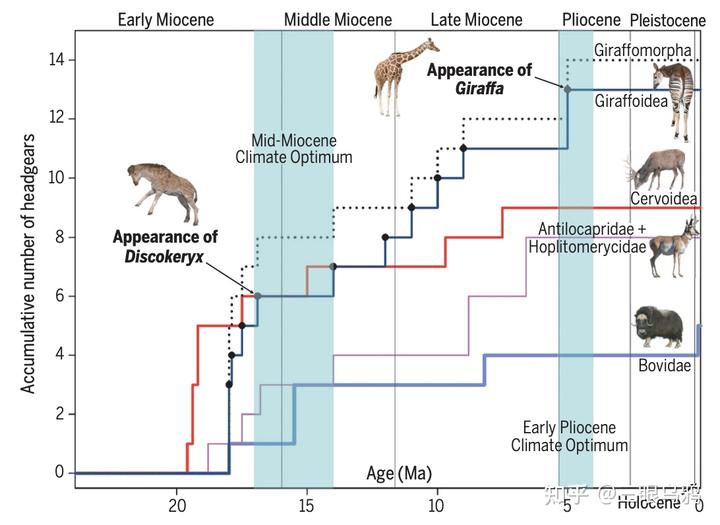
化石证据表明,长颈类头颈系统的极端演化并非始于取食高度的线性竞争,而是首先在雄性间强烈的性选择压力下被快速放大 。

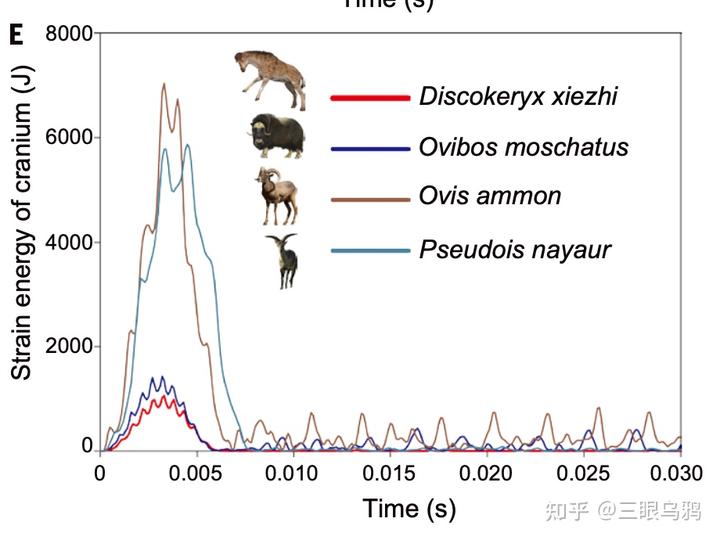
中新世早期的獬豸盘角鹿(Discokeryx xiezhi)提供了关键中间态:其颅骨、枕—寰关节及多节颈椎形成高度强化的整体力学结构,明显适应高速、正面头对头撞击。
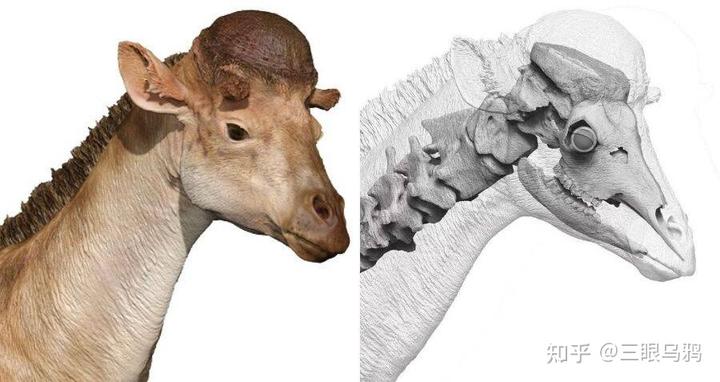
有限元分析显示,该结构在能量吸收与脑部保护方面的效率超过所有现生头撞型有蹄类,指向高频、激烈的雄性搏斗行为而非单纯的取食功能优化(Wang et al., 2022)。
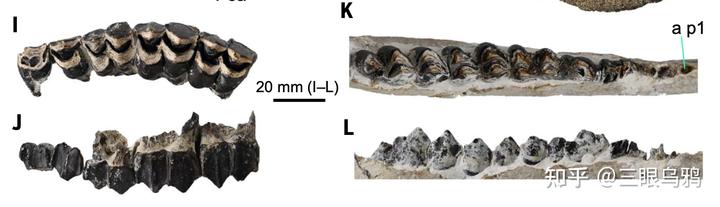
牙釉质稳定同位素结果表明,獬豸盘角鹿在部分地区显著偏向高 δ¹³C、且 δ¹⁸O 离散度较大的取值区间,指向以开阔环境为主、并利用多样或季节性水源的生境组合。
这种生态信号在哈拉玛盖等特定区域尤为突出,使其生态位与同时代牛科、鹿科明显分离,从而降低直接资源竞争,并为性选择持续、集中地作用于头—颈结构提供了稳定的环境条件。
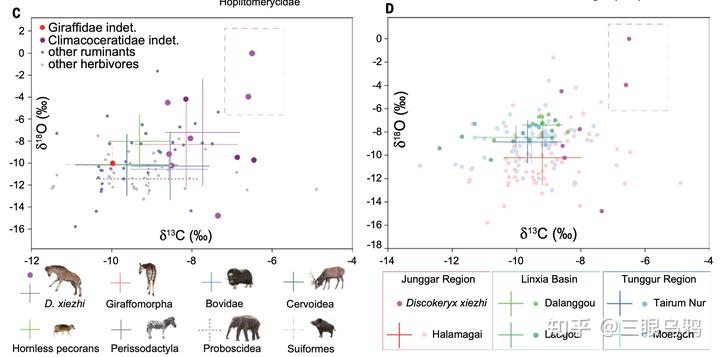
在中新世气候与生态格局变化背景下,早期长颈类通过进入边缘生态位减少与牛科、鹿科的直接竞争。在这些环境中,雄性间激烈的性选择搏斗成为主要选择力,推动头饰与头—颈系统向极端结构演化。

到上新世,现代长颈鹿的长颈—高位取食形态,才在颈搏(necking)行为基础上被进一步放大,并获得显著生态收益。

我们已经从直观的外形对比和生活习性层面讨论过不同颈部形态的差异,但仅凭外观和行为仍不足以解释这些差异为何会长期稳定存在。
从解剖结构和力学层面看,头颈系统不存在通向“最优形态”的统一演化路径,而是受不同功能需求与代价约束的共同塑造,沿着至少三条彼此分化、相互排斥的演化方向展开:
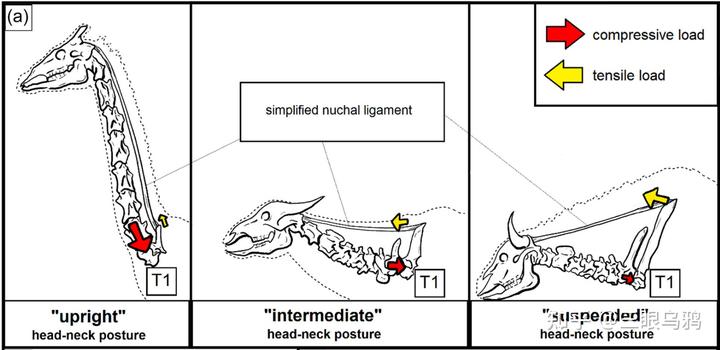
以长颈鹿、霍加狓、各类典型羚羊(如黑斑羚)以及羊驼类为代表的直立型,其颈部习惯性向上耸起,骨骼结构呈垂直堆叠。

以驼鹿(Alces alces)、双峰驼、单峰驼以及民都洛水牛(Bubalus mindorensis)为代表的中间型,其头颈高度通常与肩部持平,其中民都洛水牛虽体型较小,却表现出极高的峰值拉应力。

在自然放松或行走状态下,马类头颈高度通常与肩部持平或略高,符合中间型特征。其项韧带高度发达,从后枕延伸至胸椎棘突,使头颈在低头取食或水平奔跑时主要以悬挂方式承重,显著降低肌肉消耗。其 T1 同时承受头部牵引与颈部压缩,受力模式接近驼鹿或民都洛水牛。人工选育可使不同品种在姿态上向直立型或更水平的中间型发生偏移。

而以欧洲野牛(Bison bonasus)、爪哇野牛(Bos javanicus)、非洲水牛和角马(Connochaetes taurinus)为代表的则属于悬挂型,这类物种头部习惯性低垂,第一胸椎承受着巨大的峰值拉应力,因此演化出了高耸的棘突和强健的项韧带来支撑沉重的头颈部,形成了典型的强壮搏斗姿态。

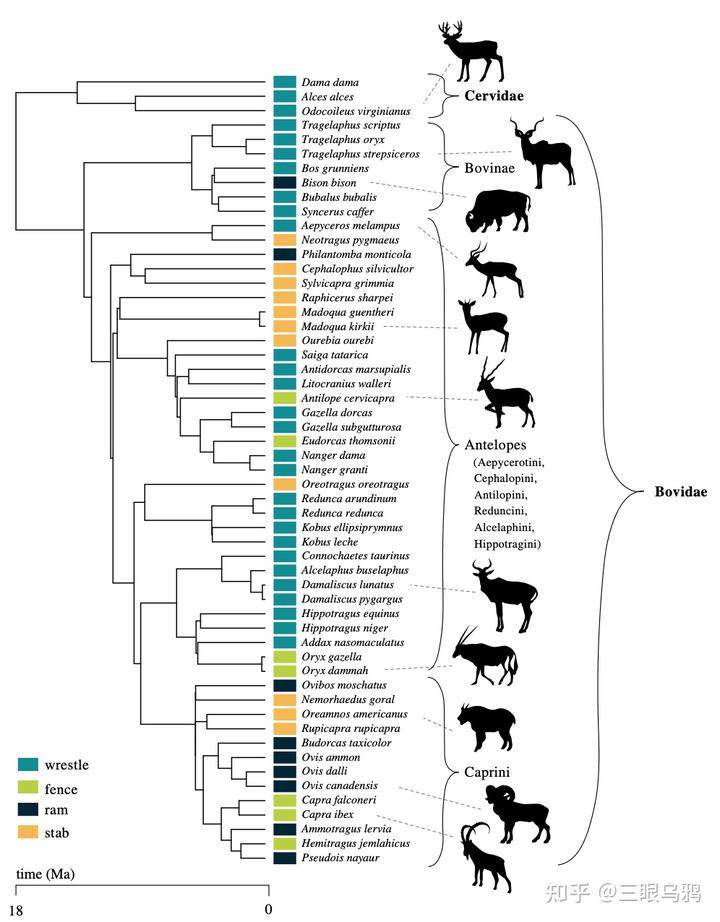
Schüler 等(2024)通过对 36 种偶蹄类第一胸椎(T1)的有限元分析清楚表明,头颈姿态与载荷类型在颈—胸交界处留下了可量化、方向一致的力学信号,这为理解不同演化路线的代价与约束提供了关键证据。
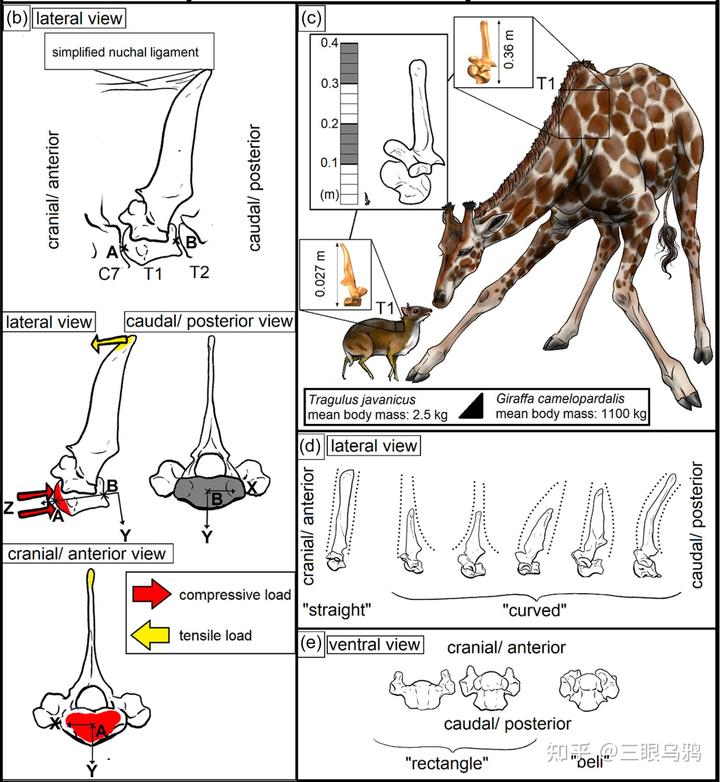

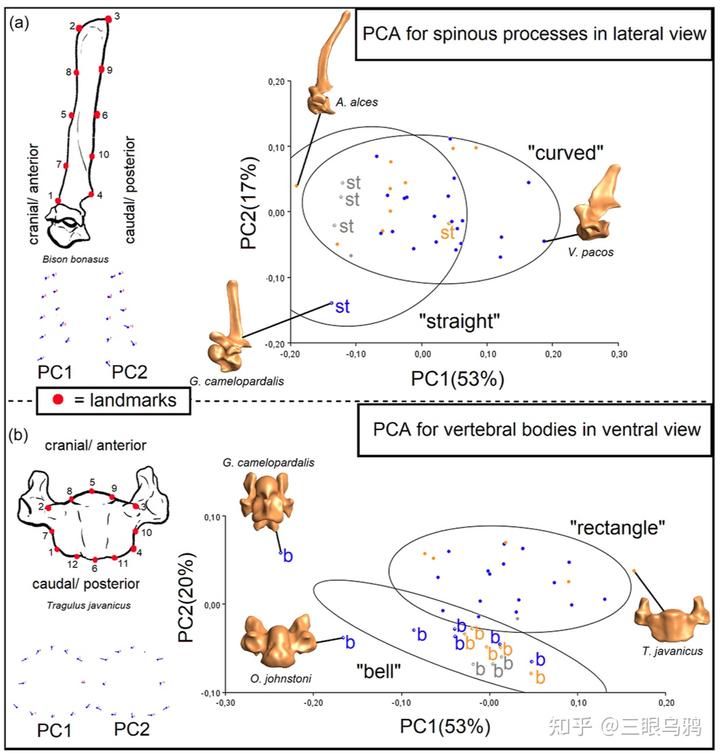
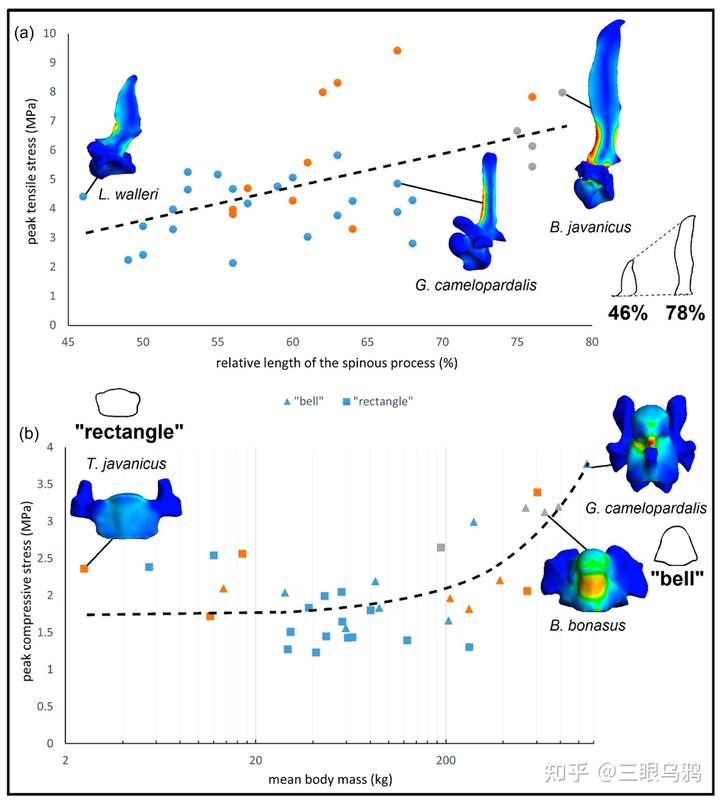
垂直型的核心特征是头颈长期处于近竖直姿态,颈部重量主要以轴向压缩的形式传递至胸椎系统。理论上,这一策略要求极强的承压能力。然而,2024 年的结果显示,长颈鹿恰恰在 T1 这一关键节点上表现出异常偏高的压缩应力。其原因并非建模误差,而是明确的形态约束:长颈鹿的 T1 发生了明显的颈化 *,椎体被拉长而非加宽,在单位面积标准化载荷下承受更高应力。
* 颈化(cervicalization)指的是原本具有典型胸椎功能的椎骨,在演化过程中在形态与力学上逐步向颈椎特征转变。
这意味着长颈鹿并未在颈—胸交界处实现传统意义上的结构加固,而是将部分承重职能后移至 T2 及更后位胸椎。由此获得的是无可替代的取食高度优势和垂直生态位独占,但代价同样清晰:T1 同时承受更高的拉伸与压缩应力。它并不是结构上的“优化”,而是用稳定性的下降,换取颈部长度与功能的扩展。
中间型则代表了一条完全不同的路线。Schüler 等的分析显示,这类动物在压缩与拉伸载荷下的 T1 应力水平居中,既未极端强化承压,也未显著削弱稳定性。2019 年的比较研究进一步补充了行为层面的解释:以驼鹿、骆驼及马类为代表的中间型动物,并不依赖高峰值正面冲撞或长时间角力,其颈椎形态在控制体重后,并未显示出针对单一载荷类型的极端偏置。

这种策略保留了较好的多方向运动能力、取食灵活性与整体稳健性,是一种典型的通用型解决方案。正因为如此,中间型的核心优势在于平衡而非极端,一旦沿长颈鹿方向显著拉长颈部,不仅会提高弯矩和压缩应力,还会破坏原有的功能均衡,得不偿失。
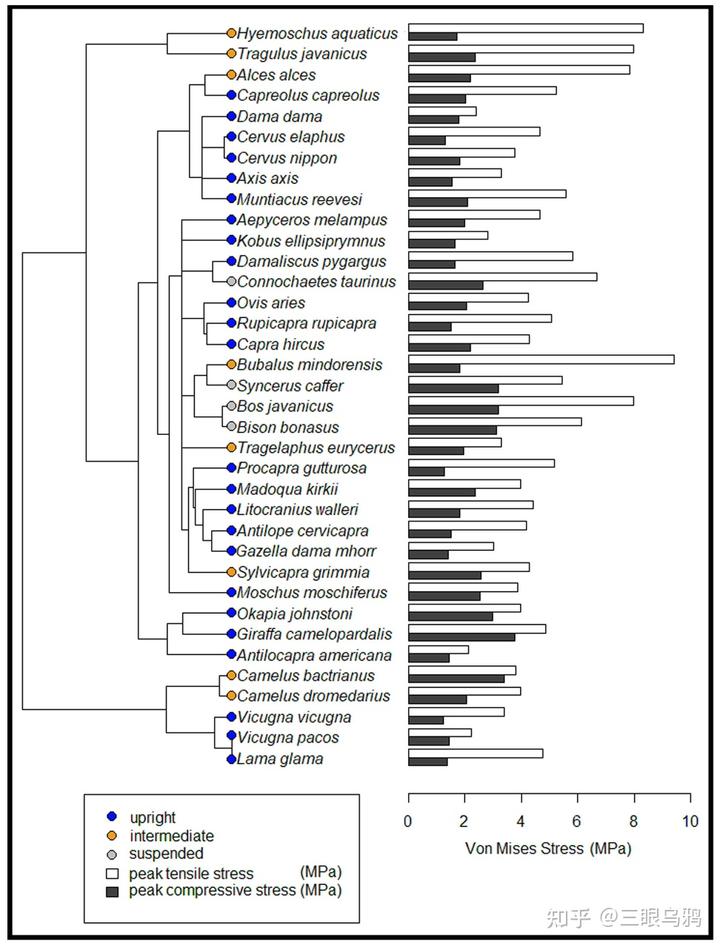
悬垂型则是第三条、同样不可替代的演化方向。以非洲水牛、各种野牛和大角羊为代表的强壮搏斗型动物,在 2019 年的研究中被明确归入以正面高速撞击和高峰值冲击为主的搏斗类型。其颈椎形态呈现出高度一致的稳定优先特征:前位颈椎短而宽,后位颈椎尤其是 C7 具备异常高的神经棘,为颈背伸肌提供巨大的力臂。

这种结构并非为了灵活运动,而是为了在极短时间内承受并反制巨大的冲击力。Schüler 等的力学结果与此高度契合:悬垂型物种在承压工况下表现出更高的 T1 应力水平,反映出其颈部更多依赖韧带与肌肉悬吊体系,而非单纯轴向承重。

对这一路线而言,稳定性本身就是核心适应目标,任何显著的颈部拉长都会放大弯矩、削弱抗冲击能力,从而直接损害其搏斗优势。

综合两篇研究可以看出,这三种头颈策略并非连续光谱上的不同点位,而是由力学需求、行为模式与形态约束共同锁定的三条演化通道。
长颈鹿的垂直化提供了巨大的生态收益,但以牺牲颈—胸交界稳定性为代价。中间型维持功能均衡,避免极端风险。悬垂型则将稳定性推至优先级顶端,以支撑高强度搏斗。
后两者若沿长颈化方向发展,必然丧失其核心优势,因此在演化上会被迅速筛除。这正是为什么长颈鹿的道路既极端又孤立:它是一条成功但代价高昂、难以复制的演化分支。

但需要强调的是,历史上并非只有长颈鹿一条谱系曾尝试过类似方向。南美洲曾存在过长颈驼(Macrauchenia patagonica,属于滑距骨目),它是中新世晚期至更新世末期的特有大型植食性哺乳动物。


它拥有相对修长的颈部和四肢,取食高度明显高于马、牛等低位啃食型动物。这表明在不同大陆,长颈化曾多次作为一种潜在解法被自然选择试探。

然而,这类形态往往意味着高度的特化,对生态环境依赖极强。在更新世末期气候剧烈波动和人类进入美洲的背景下,长颈驼未能延续,这也侧面反映了这种特化形态在面对环境动荡时的脆弱性。


从上面的例子可以看出,长颈的稀有并非偶然,而是强约束下的必然结果。马和牛的成功,建立在避开这条高风险路径之上。



而长颈鹿的存在,则是一次条件苛刻、代价高昂却在特定时空下恰好成功的稀有例子。

引用参考
Wang, S.-Q., Ye, J., Meng, J., Li, C., Costeur, L., Mennecart, B., Zhang, C., Zhang, J., Aiglstorfer, M., Wang, Y., Wu, Y., Wu, W.-Y., & Deng, T. (2022). Sexual selection promotes giraffoid head–neck evolution and ecological adaptation. Science, 376(6597), eabl8316. https://doi.org/10.1126/science.abl8316
Schüler, S., Sharp, A. C., & Nyakatura, J. A. (2024). Comparative finite element analysis of the first thoracic vertebra in artiodactyls. Journal of Morphology, 285, e21695. https://doi.org/10.1002/jmor.21695
Vander Linden, A., & Dumont, E. R. (2019). Intraspecific male combat behaviour predicts morphology of cervical vertebrae in ruminant mammals. Proceedings of the Royal Society B: Biological Sciences, 286(1911), 20192199. https://doi.org/10.1098/rspb.2019.2199