

大脑的结构中,树突轴突、突触、离子输运等等结构,是大脑功能的基础。这些结构一般在 1 到十几个纳米范围内,它们肯定是基于量子力学机制才能发挥作用的。所以说,人脑的运转中当然有量子力学机制。
但是,这是人脑逻辑部件的硬件机制,和题主的问题是两码事。就好像说,我们问类似的问题:
“电脑运转到底有没有量子力学机制?”
我们说,当然有。集成电路、芯片、逻辑门、乃至底层的半导体能带结构,这些都是量子力学机制。没有量子力学,我们造不出现在的硬件,也就不可能有一个运转的电脑。但是我们肯定不能说,因此我们的电脑就是量子计算机。因为在软件层面上,整个运算逻辑都是经典的。
所以,现在的问题就是,抛开人脑的硬件机制中的量子力学效应,在逻辑运算和信息处理上,人脑是经典的还是量子的?
我这里搬运一篇文章的观点,来自 Max Tegmark[1]。在他看来,由于退相干时间的极其迅速,以及神经元激发和传导的过程极其缓慢(相对而言),人脑的运算逻辑中不太可能有量子机制。
这里就涉及到了我们对神经激发和传导过程中的退相干时间的估算。这里的研究对象是神经元细胞中最关键的部分,轴突。我们知道,轴突可以看作是一条长长的中空纤维管,直径大于是十几微米,而管壁的厚度大约是十几个纳米。整个轴突被绝缘的髓鞘包裹(可以想象成为电线外面的绝缘包皮),但是这个绝缘层每隔一段距离就中断一个空隙(兰氏结),裸漏出内层的管壁。如下图所示:
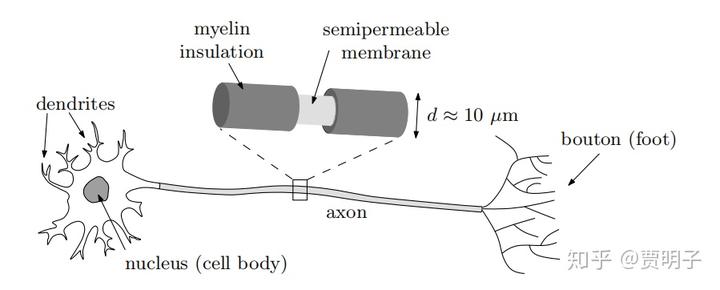
裸漏出来的管壁是一个对各种离子的半透膜,由于离子的电性,所以内外日常维持着一些微妙的电位平衡(具体细节不在本文范围内略过不谈)。当神经元在激发过程的时候,由于电位的刺激,会打开半透膜上的电压钠离子通道,瞬间大量的钠离子涌入内部,然后沿着轴突传导,再激发下一个兰氏结处离子通道的开关状态,完成电信号沿着神经元的传导。这个传导速度大约是 100m/s。每次激发的时间大约是几毫秒,然后钠离子通道关闭,神经元渐渐恢复原来的电位。
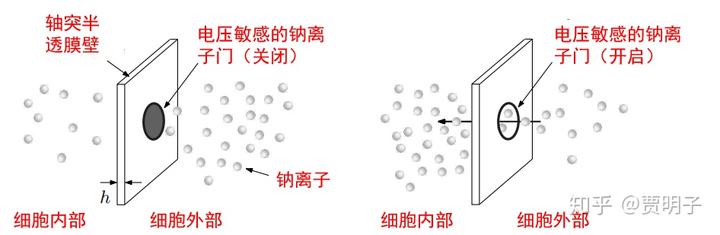
通过神经科学的实验发现,Tegmark 估算,再一次神经冲动过程中,大约有 10^6 个钠离子完成了这次传输过程。也就是说,从量子力学上,神经元的激发状态和静息状态的叠加态,就是这 10^6 个钠离子的整体状态的叠加态:
这些离子与外界环境密切相互作用。包括了与周边的钠离子、水分子、以及与周边的电场的相互作用。这种系统与环境的密切耦合就必然导致退相干。Tegmark 用粒子散射模型进行了估算,大约算出,退相干时间在
。也就是说,这是维持叠加态的时间尺度。
然而,神经元的一次激发过程大约为毫秒级,Tegmark 进而估算,人类的大概认知过程(如思考、精细动作、感知)的时间尺度大约是零点几秒。也就是说,相比于人类的意识活动,退相干基本上可以看作是瞬时的。整个神经网络的激发过程,就是经典过程。
此外,Tegmark 还估算了神经元中的神经微管的退相干过程。神经微管是神经元中的关键结构,曾被某些人认为是最有可能形成量子计算的地方,甚至被认为是“人类意识的起源之处”[2]。
神经微管是由 13 根所谓的原纤维拼接而成的,每根原纤维则是由一系列的“蛋白二聚体”(tubulin dimer,百度翻译而来,不知准确否)连接而成。整个微管直径 24 纳米,长度可达到微米到毫米级。每个 tubulin dimer 由一个α单体和一个β单体构成,带有明显极性。拼接成长纤维的时候,就形成了两段的电位差。
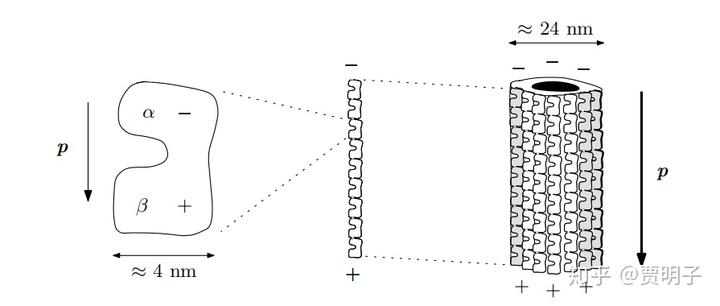
而这中间有一个相对自由的电子,它的状态可以引发整个 tubulin dimer 的两种不同状态,对应不同的极性(就好像自旋态那样)。因此它的变化会导致极性沿着整个纤维快速传递。
Tegmark 以周边钙离子的电场干扰对一长段纤维进行估算,估算的结果大致上退相干时间为 10^-13s。这个时间要比前面提到的激发态时间长得多,但是仍然,比起人类意识而言,也可以算作瞬间了。因此,它参与量子计算的可能性也微乎其微。
Hagan[2]等人对 Tegmark 的结果进行了挑战,他们计算单个的 tubulin dimer 而不是一段纤维的退相干时间,认为,单个 dimer 的退相干时间要比整个纤维长若干数量级,因此完全有可能形成较长的量子相干态。但是他们的结果又被 Rosa 等人质疑[3],认为他们的一个基本假设有误,而实际上退相干时间和 Tegmark 估算的大致相似。
总而言之,现在看来,还没有证据显示大脑内有量子计算的迹象。