


正是因为我们祖先守住了五根手指,我们才逆袭成为灵长之长。
当你无意中看到自己的五根手指,你可能想不到,决定你五根手指的基因来自 3.5 亿年前的古老石炭纪。
如果你的手指是一个单独的物种,它就是典型的活化石。
4.19 亿年前,梦幻鬼鱼[1](Guiyu oneiros)在水里自由自在游弋时,恐怕不会想到,它们其中一支后裔会发展出四肢和指 / 趾骨,征服它们从来没有踏足过的大陆,创造一段又一段演化传奇。
梦幻鬼鱼是最早的硬骨鱼之一,同时也处在肉鳍鱼演化树的基干位置(最底部)。
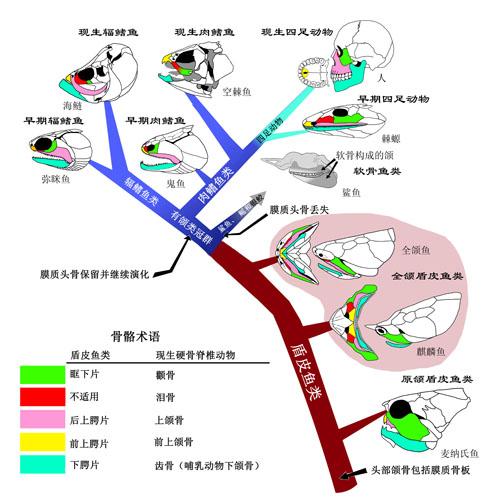
我们的上肢和下肢正是来源于它们的胸鳍和腹鳍。
可以看出,鱼鳍的主要结构便是鳍条。
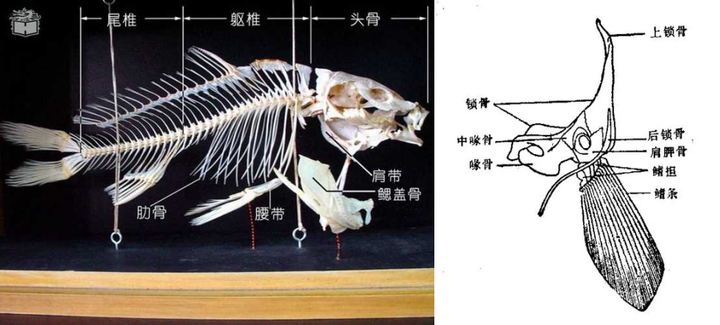
很多人可能会理所当然地认为:
人类的手指,应该是由鱼的鳍条演化而来,或者是一种全新发展出来的结构。
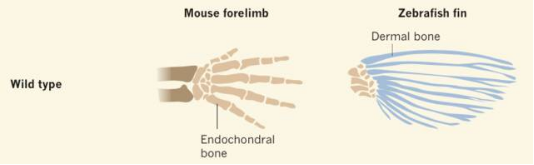
其实早期生物学家也这么想,但却被后来的研究打了脸[2][3][4]。
组成鳍条的膜骨,本质是一种结缔组织,与软骨内成骨的指 / 趾骨并非同源结构。
所以,指 / 趾头不可能由鳍条发育而来,而是来源于鳍条基部的软骨内成骨。也就是鳍条和肩带 / 腰带间连接的很小一部分,看起来的确很不起眼。站在 4 亿年前来看,谁能想到这么小小的一部分,在未来能够演化到足足可以占到整个体重的 50%呢?
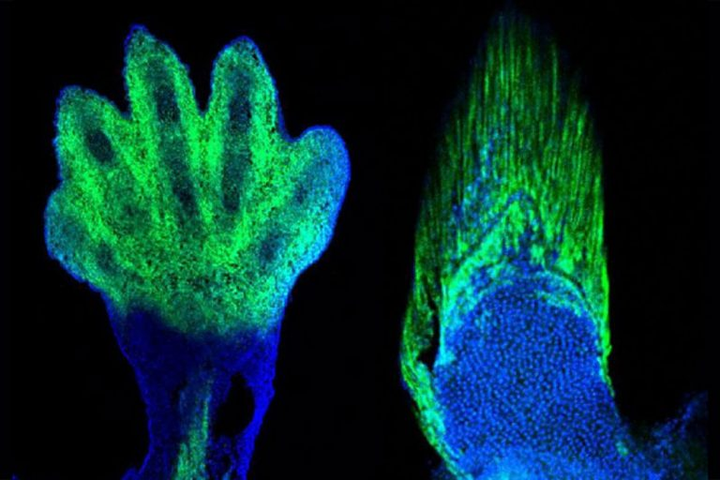
软骨内成骨是否发展出指 / 趾头,由 Hox13 基因家族所控制。
- Hox13 基因并不是单个基因,而是分布于不同染色体上的多个基因。
2016 年发表于《nature》的研究发现,如果敲除斑马鱼的相关基因(hoxa13a、hoxa13b、hoxd13a),它们的鳍条就会发育得非常短。甚至随着鳍条缩短,基部会出现了更多的软骨内成骨,这和组成指头的骨骼是相同的类型。相反,如果敲除小鼠的相关基因(Hoxa13、Hoxd13),它们便无法发育出手部结构。
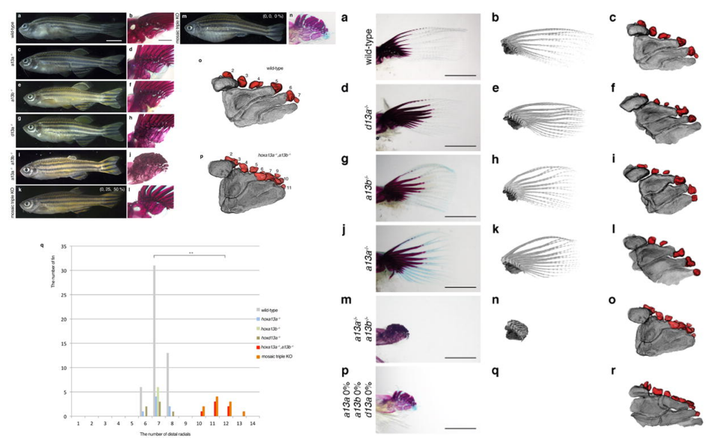
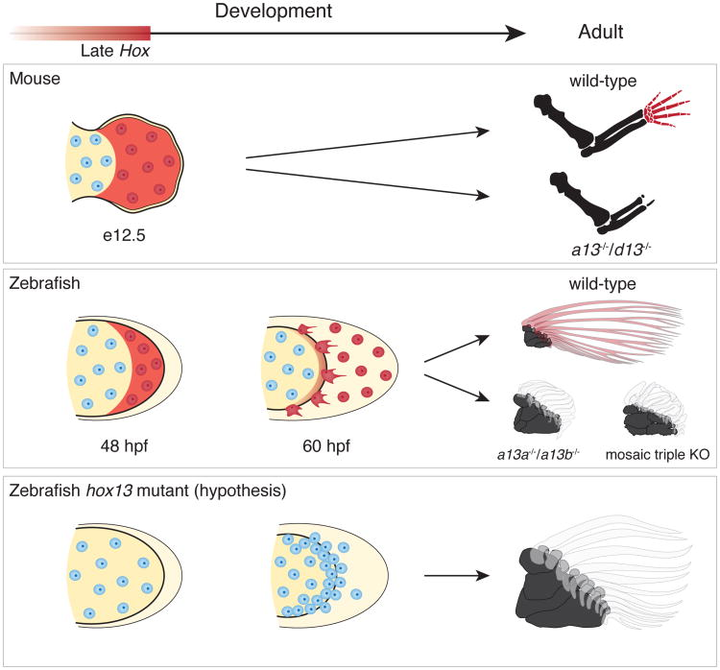
从这个实验我们可以推测,在肉鳍鱼演化的过程,大概率正是通过关闭 / 开启相关的 Hox13 基因,来指导完成从鳍条发育到指骨发育的转变。时至今天,偶尔出现多指症的人类,正是与 HOX13 相关基因的突变有关[5]。
演化本身就是一场修修补补,遵守最大化利用原则。
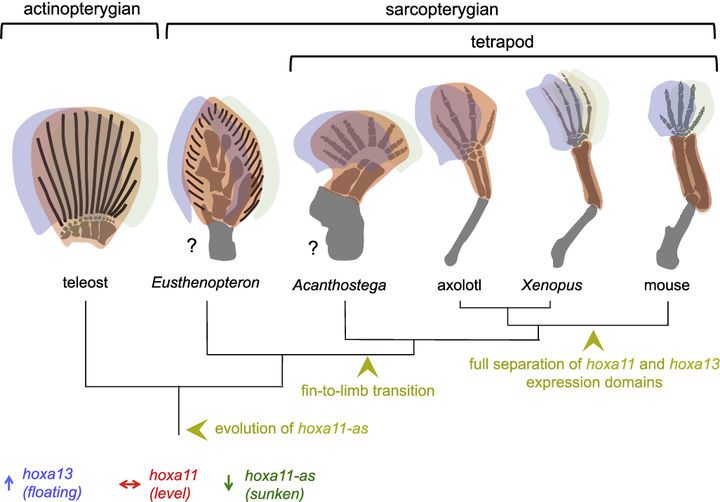
4.19 亿年前,梦幻鬼鱼鱼鳍基部的那一点点软骨内成骨基质,经过 3000 万年的时间,逐渐发展出拥有四肢骨、腕骨的一大类肉鳍鱼,以 3.9 亿年前的真掌鳍鱼[6](Eusthenopteron)为代表,它们已经开始朝着陆地进军。
然而,它们最终以失败惨淡收场。

除了四肢还不够强壮外,它们失败有一个重要的原因,就是没有演化出指 / 趾骨。
又过了 1000 万年,3.8 亿年前的潘氏鱼[7](Panderichthys)总结失败的教训,发展出骨化程度很高的肱骨、尺骨以及桡骨,尤其是肱骨,比同时期的其他任何鱼都要长。再加上拥有强壮的肩胛骨和骨盆带,它们可以在浅滩上进行足够长距离的爬行。
为了适应浅滩爬行,不知经历过多少次匍匐前行,它们终于发展出了趾骨。
虽然最开始,指 / 趾骨更像是一些散碎的骨头,但通过指 / 趾骨支撑出更大的面积,不仅让它们在浅滩爬行更有力量,还可以通过蹼相连,同时保持优秀的游泳能力。

然而生物演化最忌讳的就是中间生态位,高不成低不就,潘氏鱼最终也抱憾而终。不过与它们亲缘关系较近的一支却越挫越勇,不断改造指 / 趾骨,先后发展出提塔利克鱼(Tiktaalik)、希望螈(Elpistostege),并最终在 3.65 亿年前左右,演化出了具有典型指 / 趾骨的棘螈(Acanthostega)、鱼石螈(Ichthyostega),以及图拉螈(Tulerpeton)[8][9]。
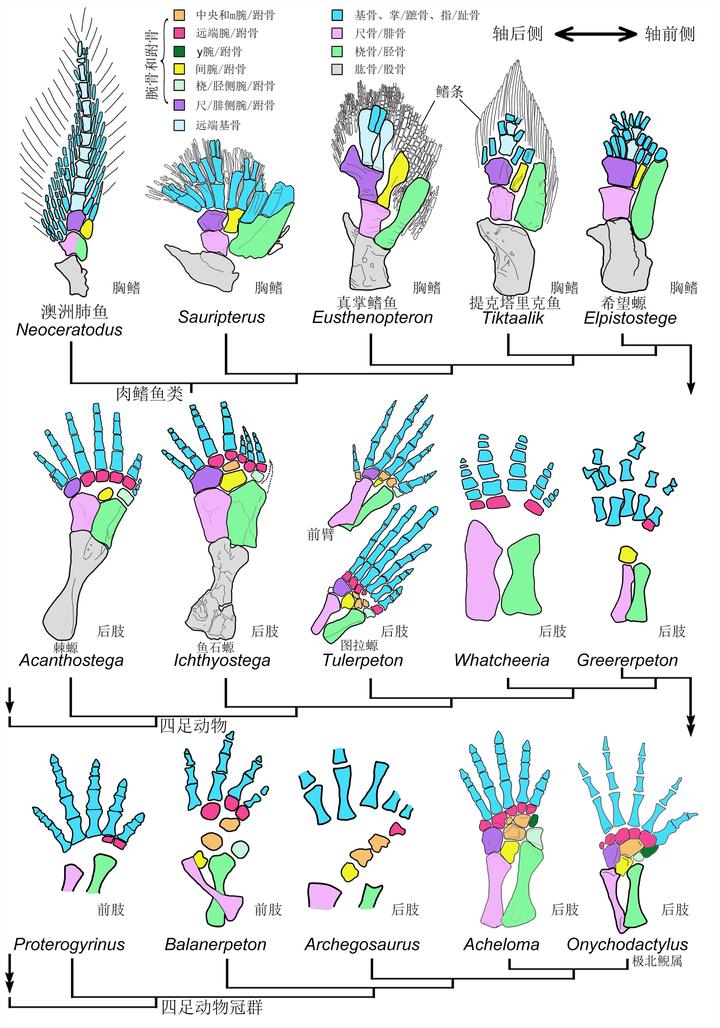
棘螈足足有着 8 跟指 / 趾骨,不过以中间六根为主。鱼石螈则丢失了一根,为 7 跟指 / 趾骨。到图拉螈时,则已经只剩下 6 根指 / 趾骨。
时间到了 3.48 亿年前,出现了一种代表生物——彼得普斯螈(Pederpes)[10]。

后趾 5 跟,前趾 6 跟,被认为是功能上的五趾动物。
从此以后,早期四足动物逐渐从多指型,发展为典型的五指型。
不过早期四足动物的五指型可能并不是单起源,而是多起源的。
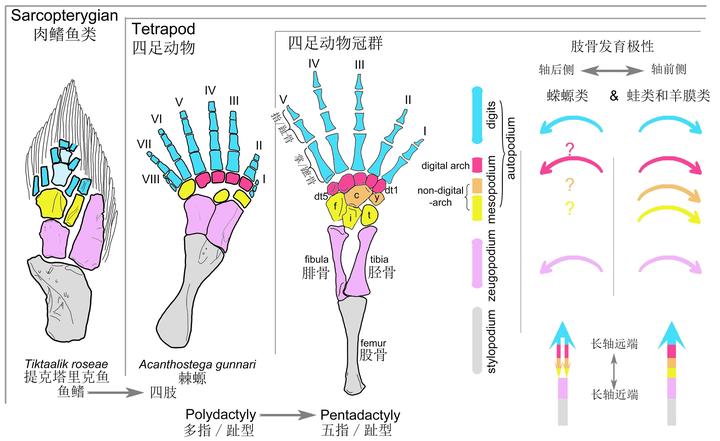
现今的所有四足动物,都是由其中一支五指型发展而来的。
其它所有指型,全都在四足动物登陆后不久,全部灭绝了。
在水环境中,鱼类前进,主要面对水的阻力,优化身体的流体力学结构就可以了。但在登陆后,它们的身体和四肢,必须前所未有地抵抗重力。因此,无论四肢、掌骨、还是趾骨,都随着陆地爬行产生的应力,发生全新的适应性变化。

在重力下,它们腕骨(前肢)和跗骨(后肢),发生了轴向前折。身体重量全部施加在腕骨和跗骨位置,造成骨骼发生重新排列。
手掌内部和外部两块小骨的内缩,使得指骨附着面积减少,最终造成了边缘指 / 趾骨的退化,手指数目自然便逐渐减少。
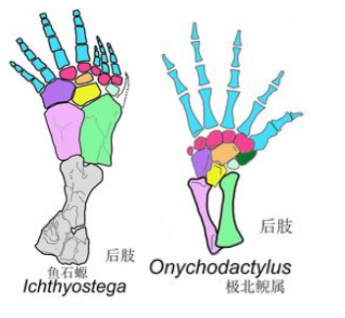
除了手指减少的第一演化压力,来自腕部的力量支撑需要外,更少的指骨也可以避免过多手指间不必要的竞争,可使手掌上附着的肌肉更加集中,使得双手变得更加灵活有力。因此,四足动物登陆后不久,便逐渐掌握了抓握和攀援能力,从而可以迅速辐射发展到各界各地复杂的生态位中去,遍及山川湖泊。
你可能会有疑问,那为什么手指没有减少到 4 根以下,而是保持在 5 根的数目呢?
主要原因在于早期四足动物大多都是两栖动物,它们同时还有着游泳需求。很明显 5 根手指相起比更少的手指,可以支撑起更大的面积,从而有利于游泳。另外, 它们的骨骼的强度较低,支撑身体的指骨也不能太少。
虽然五根手指是四足动物比较原始的特征,但却具有朝着各个方向发展的非凡潜力。
生物演化的魅力在于,原始并不等于失败。失败的唯一标准,是被大自然从地球生态中彻底抹除。
随着脊椎动物向大陆全面征程,原始的五指 / 趾,终将随着一次次王朝兴衰,发生全新的惊人变化。
陆生脊椎动物行进的方式主要分成跖行、趾行,以及蹄行。
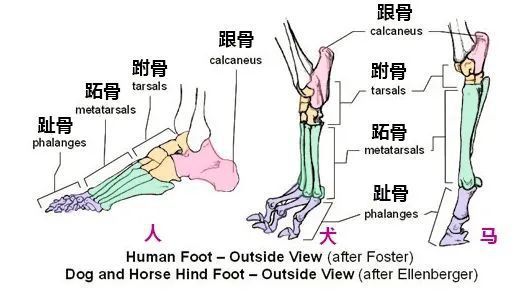
最早的四足动物全部都是跖行,至少长达 1 亿年的时间,步态都没有发生巨大的变化。最主要原因在于它们基本都是冷血动物,新陈代谢能力有限,它们要么奔跑速度不够快,要么不能长时间持续奔跑。
时间到了 2.5 亿年前的二叠纪末期,随着中温动物乃至恒温动物的出现,动物的活动能力增加。不仅速度慢的植食动物会被吃掉,速度慢的掠食者也会被饿死。
有跑步经验的人应该知道,高速奔跑时,抬起脚后跟,脚趾抓地,接近趾行的状态,能奔跑得更快。随着速度演化越来越内卷,不能保持高速步态的动物灭绝,最终活下来的步态发生了彻底改变,成了完全的趾行动物。一开始的趾行动物,也都全部具有五根指 / 趾头。
不过指 / 趾头越多,意味着力量更加的分散,对奔跑速度也越有影像。
因此,随着速度需求的增强,指 / 趾头就会具有退化趋势。不过对于有着抓握和攀爬需求的动物来说,又具有反向的演化压力来对抗趾头的减少。
例如,今天的猫科动物,前爪还具有抓握和攀爬需求,因此保留了五爪。后肢抓握需求降低,更加倾向于奔跑,因此只有四趾。
不过最早出现趾头减少的动物,并不是哺乳动物,而是恐龙。
二叠纪末期大灭绝事件之后,随着合弓纲动物的衰退,主龙类崛起。从 2.3 亿年前的三叠纪晚期开始,恐龙足足统治了地球一亿多年,它们内部也是风起云涌,王朝更替,期间不同分支发生了不同步态的变化。除了牢牢占据巨型生态位的原始分化类群——蜥脚类是五趾外,后来的类群或多或少都经历了高度的进化竞争,大多数恐龙都退化了脚趾,成了四趾动物,也有的成了三趾动物,甚至有的只有两趾,以恐爪龙为代表。
当然由于中晚期恐龙前肢需求大大降低,严重退化,甚至出现了一根手指的单指临河爪龙。

指 / 趾头的变化,体现了一个物种的生存竞争压力。它们不同形态的指 / 趾形,折射出的是它们曾经的辉煌。今天的鸟类依旧存在不同的趾形,同样与奔跑、抓握等不同生境需求息息相关。
我们的祖先,合弓纲这一支,经历二叠纪末期的大灭绝,三叠纪的大干旱的,最终只有一些小型动物演化成了哺乳动物。为了躲避恐龙的锋芒,它们钻进丛林里成为夜行动物,才博得了一线生机。
生态位的长期稳定,也使得它们的演化异常缓慢。
最早的哺乳形态类动物出现于 2.25 亿年前,以隐王兽为代表,几乎和恐龙同时出现。然而一直经历了近 6000 万年的时间,到 1.6 亿年前,它们才发展出了最早的胎生真兽类动物——中华侏罗兽。而真兽则在接下来长达近 1 亿年的时间里,无论体型还是生态位都没有发生多大的变化。

在夜行丛林生态位上,我们祖先不仅有攀爬的需求,挖洞的需求,也有捕捉昆虫的需求,采摘果实需求,这些都我们的双手一直保持着保守的原始特征——五指状态。
6500 万年前,恐龙灭绝后,整片大陆再次留下巨大的生态位真空。
不同的哺乳动物适应不同的生态位,再次发生了一场场特殊的进化竞争,四肢再次发生了巨大的变化。尤其以有蹄类这一支最为显著,不仅演化成了今天偶蹄目和奇蹄目,一支甚至索性再次下海,演化出了鲸类,完全退化了后肢。
然而,我们祖先灵长类这一支,任它几经沧海桑田,承担多少兴衰荣辱,终究没有选择退化五指,而是爬上树梢,不断强化手指的抓握能力和灵活性,随后又走出丛林,不断锤炼我们的双手。使它成为地球上最强的攻击性器官,最终缔造了属于我们自己的辉煌。
当然,这又是下一个故事了。从某种意义上来说,正是我们祖先把原始的五指特征坚守了 3.5 亿年的时间,才让我们今天有机会创造出人类王朝。