


当然是……
会
吸血吸得爽到爆~
爆炸:

不过,这并不会在正常蚊子身上发生。
上图中的蚊子是被处理过的。
虽然正常的蚊子,一次性吸血可以超过自身 3 倍体重,乃至于清晨你看到那原本纤瘦灵活的蚊子,变成了圆滚滚的带翅膀的笨重小血球,可以被你轻松拿捏爆浆……

但它们正常的生理机制,并不会蠢到会无限吸血到爆体而亡。
当雌性蚊子吸血达到 2~3 倍体重时,随着腹部的机械张力增大,它们位于腹神经索上的的腹部伸展受体 / 感受器(abdomen Stretch receptors)便会发出信号,终止进食[1]。
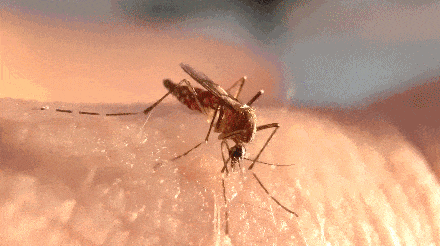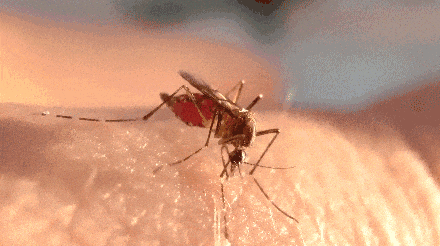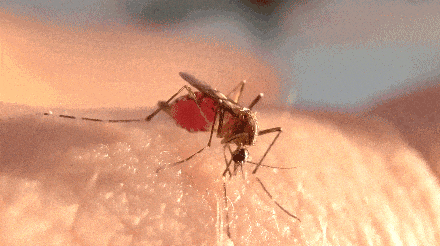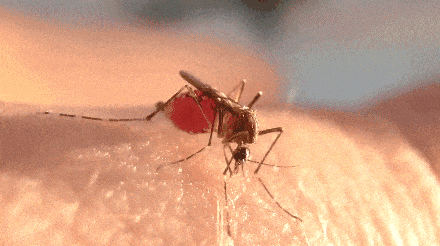
不过,题主问的却是一直处于吸血状态。
虽然它们自己不会这样,但显然可以帮它们处于这样的状态:吸血吸到爽到爆。
热爱蚊子实验的墨尔本大学研究者佩兰·罗斯(Perran Ross)做了一个这样的实验。

他直接给蚊子腹神经索前来了个高位切断,切断 / 粉碎腹神经索,破坏信号通路。
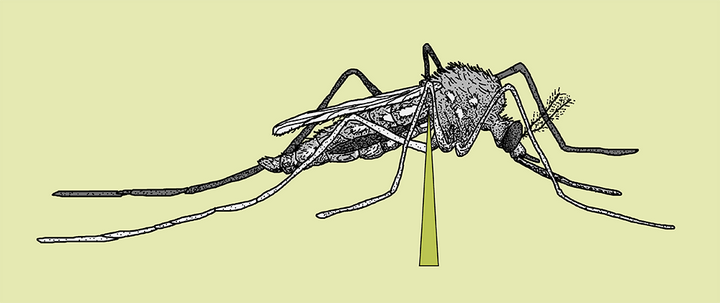
腹部的信号,无法再传递到头部。
蚊子开启了无限吸血的爽爆模式。
很快,蚊子的肚子就变得圆滚滚了起来,超过正常吸饱血的蚊子。
- 这是剪断腹神经索异常吸血的埃及伊蚊(左)与典型饱腹蚊子(右)的对比:

腹神经索被剪断的蚊子,吸血量可以轻松达到 4 倍体重,甚至最终爆体而亡。
而且腹神经索的切口越靠前,吸食的血量就越大。
这表明,蚊子的腹部伸展受体存在于整个腹神经索上面,尤其是前胃。
对了,佩兰·罗斯还有个实验壮举,就是——以身饲蚊。
他本身做的研究是利用沃尔巴克氏菌(Wolbachia)[2][3]抑制蚊子繁殖,控制登革热传播。
为了让自己的实验蚊大量繁殖,顺利产卵,他用自己的血液来供养 15000 只蚊子吸血。
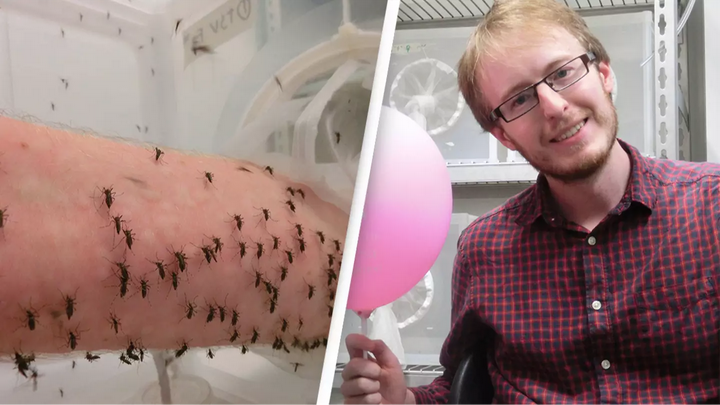
而他实验的被撑爆的蚊子,正是源于他自己的血液。
随着蚊子一代代的繁殖,我们也在佩兰·罗斯身上看到了岁月的痕迹。

看到了他逐渐拥有了强者的发型。

其实,切断蚊子腹部伸展受体,并不是佩兰·罗斯首创。
至少早在 1969 年,就已经有人做过了系统的实验[4],并并发现,剪断腹部第二神经节之前的位置,几乎所有雌蚊都会吸血到爆体而亡。
可能你依旧会有疑问,为什么完全断绝了腹部信号,蚊子就几乎都会爆体而亡?
人体具有血压,可能很多人会理所当然地认为,蚊子吸血主要是通过血压。
其实并不是。
蚊子吸食的是静脉血。
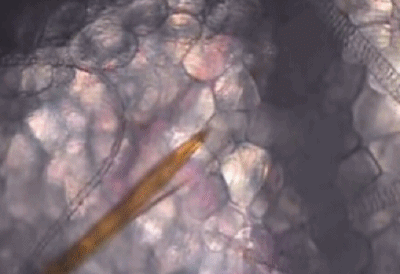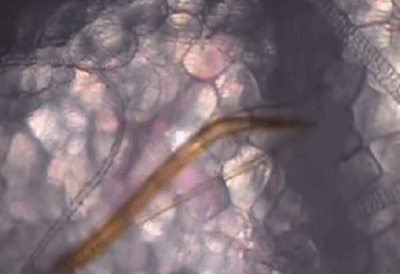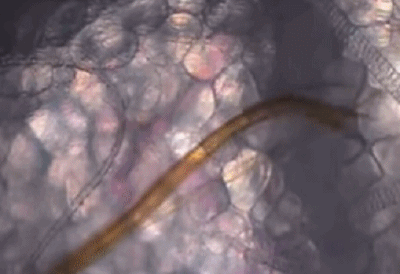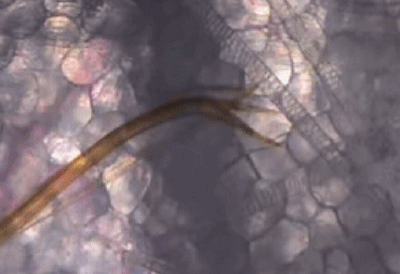
它们口器中的上唇和舌,可以在皮肤组织间不断地探索,通过特殊的嗅觉通路[5],快速找到外周浅静脉血。
蚊子吸血并不单纯依赖人体血压,而是主动提供负压吸血。
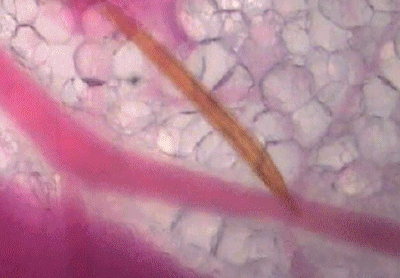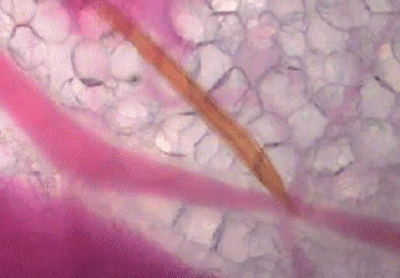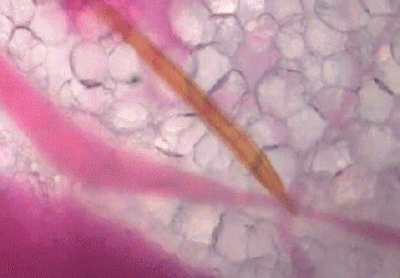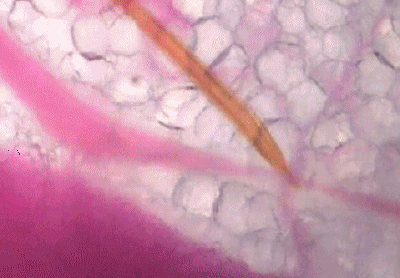
可以看出,浅静脉血管被蚊子吸到临时干瘪,具有明显的主动吸血过程。
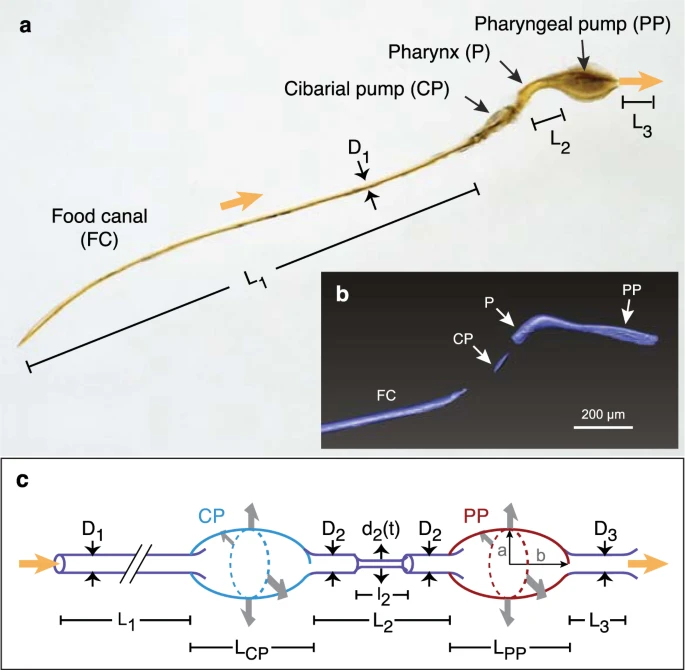
蚊子的头部具有两套吸泵:
食窦泵(cibarial pump,cp)以及咽泵(pharyngeal pump,pp)[6]。
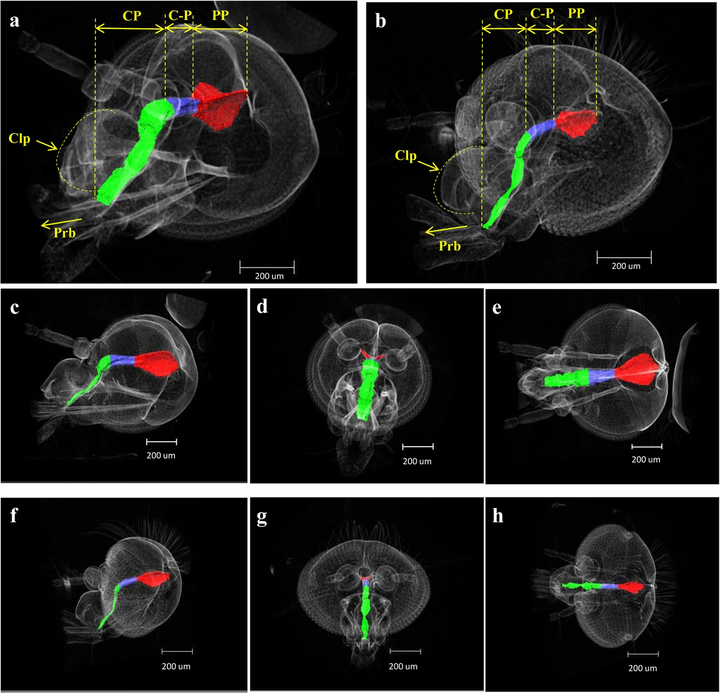
这是中华按蚊(An.sinensis)与东乡伊蚊(Ae.togoi)的两套吸泵对比。
- 绿色(cp)为食窦泵。
- 蓝色(c-p)为两个泵之间的阀门,可防止吸食时的液体回流。
- 红色(pp)为咽泵。
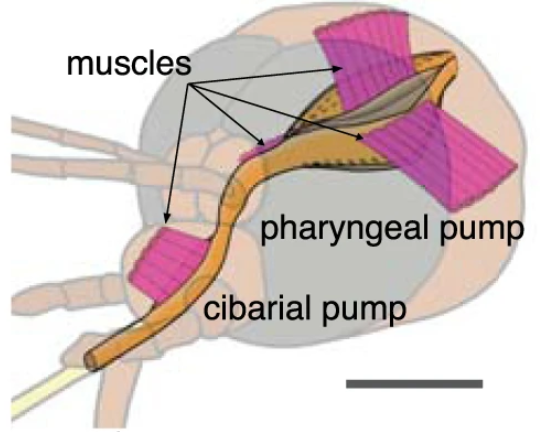
伴随着吸泵上的肌肉收缩,蚊子吸血时,两套吸泵会在短短 1 秒内完成数次交替膨胀和收缩,从而维持持续性吸血的负压。
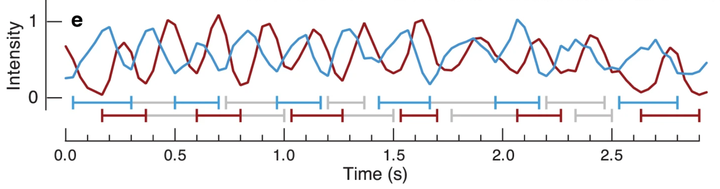
通常,蚊子吸血时提供的负压并不高,只有 0.76KPa,不超过 1KPa。
人上肢浅静脉血压大约在 1KPa 左右,即便考虑血压,提供的总吸力也大约在 2KPa 左右。
不过,蚊子在吸血过程中,还存在爆发式泵吸模式。
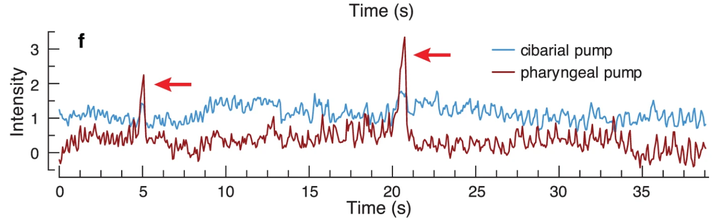
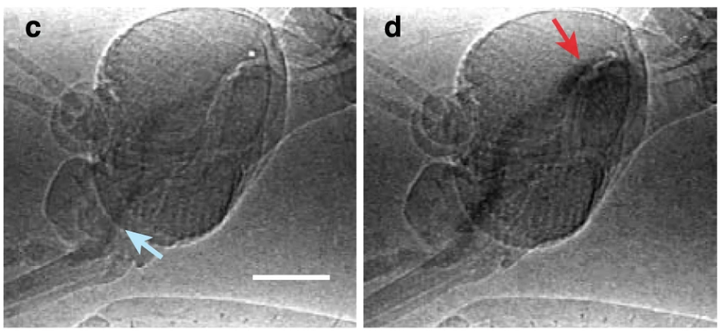
爆发模式产生的负压高达 40KPa[7](相当于大气压 / 真空负压的 40%),可以观察到明显的泵腔体积变化。
蚊子的腹部可以近似看做薄壳墙体。
对于薄壳圆管来说,压强公式为:
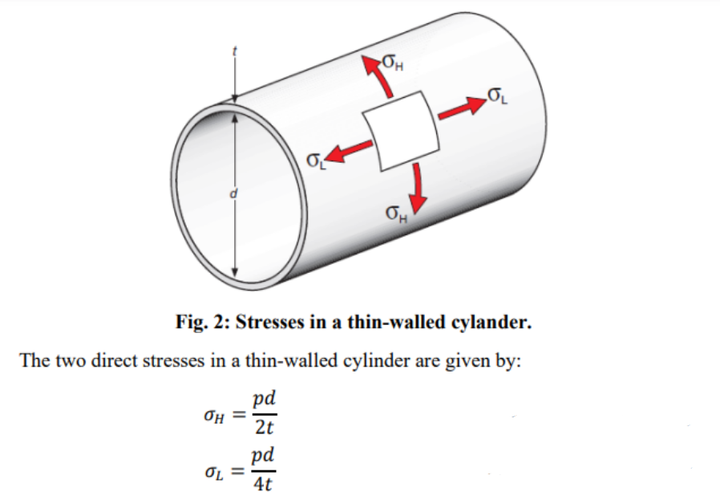
蚊子腹部几丁质厚度大约 2μm,也即 2×10^-6m。
腹部吸满血直径可达 2mm,也即 2×10^-3m。
可得,蚊子腹部受到的环向应力σH 为:
σH=500P。
可以计算出,在连续吸血模式下,环向应力为 1MPa 左右。
而在爆发吸血模式下,环向应力则高达 20MPa。
蚊子腹腔壁属于典型的柔软几丁质,且分成略微偏硬的腹节膜片,以及具有高度膨胀与伸展性的柔软链接。
膜片的强度可能超过 10MPa,但中间柔软连接的强度可能不到 1MPa。
显然,蚊子爆体而亡的位置,在柔软连接处。
可以看出,哪怕蚊子吸血不用爆发模式,仅仅连续吸血,在没有伸展受体进行信号阻止的情况下,腹部承受的应力会正好达到破裂的临界值,从而可能出现爆体而亡。
而爆发模式下,则是必然会出现爆体而亡。
总之,腹神经索被高位剪断的蚊子,会出现一直吸血的现象,而它们的吸力和材料力学属性决定了,最终大概率会吸血到爆体而亡。
PS:
蚊子的主动吸血模式,决定了,正常的蚊子哪怕吸血时口器被卡在人的皮肤中,也不会爆体而亡。人体浅静脉的血压,通常达不到让蚊子爆体而亡的地步。而且,蚊子口器存在阀门,如果不打开,人体血液也无法进入蚊子体内。
所以,正常蚊子口器卡主爆体而亡的说法,仅仅只是虚假的都市传说。