


宇宙间有光,而地球上有着追光的生命。
每天,太阳都向四周发射着光芒。其中的二十二亿分之一,以被地球人称为“光子”的形式穿过长长 1.5 亿千米的幽深空间,在地球上播撒下宇宙的辉煌。

在人类可见的其他星球,光照射在光秃秃的地表上,掀不起一丝波澜;但在地球上,日光不仅加热了浓密的大气和浩瀚的水体,搅动着和风与惊涛;更有另一小部分被生命所截获,当世无双地转化为构成有机质所需的能量。这些能量化为一株株植物,编制成一层薄薄的绿色地毯,为地球铺上翠绿的屏障。它们吸收着阳光,涵养着水源,将无穷无尽的氧气和食物提供给地球上的其他生命,并在数十亿年的演化中演化出无穷无尽的艺术和想象。
从湿滑的蓝藻,

到挺立的莱尼蕨

高耸的鳞木

长青的松柏

再到丰收的麦浪

一种处于特殊状态的叶绿素 a,暗示了我们所有光合生物的共祖性。这种叶绿素及其辅助分子被称为“反应中心”,具有光化学活性,能将光能转化成化学能。所有光合生命都具有相似的“反应中心”。以及相似的蛋白质和辅助因子结构。这意味着,它们从一个共同的祖先进化而来,并在水上陆上演化成长林水草,参天石松。

花团锦簇、稻麦飘香。我们对这一切都已习以为常。但第一个光合生命究竟来自何方?
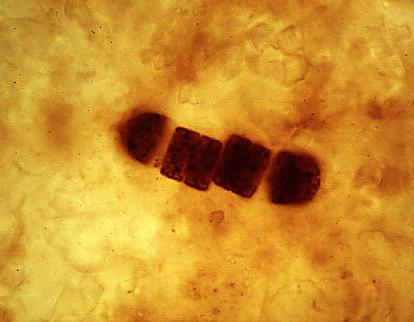
最早的光合细胞早已消失在元古代的波浪之中,但它的后代迅速改变了世界。38 亿年前的地层中,最早的生物固碳证据——改变的沉积岩同位素组成便已出现,而最早的光合生命化石,一些丝状的细胞结构,可以追溯到 35 亿年前产生。这些细胞最先被认为是释放氧气的蓝藻,但后来根据化学分析发现,这些化石更可能是光合细菌,一种在不产生氧气的情况下进行光合作用的生命。
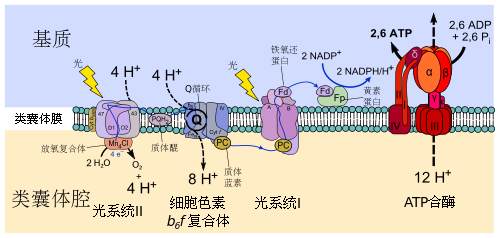
光合作用产生的氧,源自水的分解。陆生植物、绿藻和蓝藻同时使用两种反应中心:RCI(I 型反应中心)和 RCII(II 型反应中心)。这两种光系统一起吸收光能,将水分解成氢离子和氧气,产生氧气时放出的电子随即被酶接收,将光能转变为有机物中的能量,参与接下来的反应。
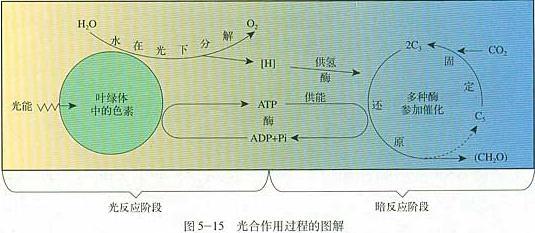
这个反应中,水只起到提供电子的作用。而水一种是非常稳定的物质,只有当酶有了非常强的氧化能力,才能从水中提取电子;那么,如果有更易释放电子的物质存在,光合生物也乐意接收并使用。
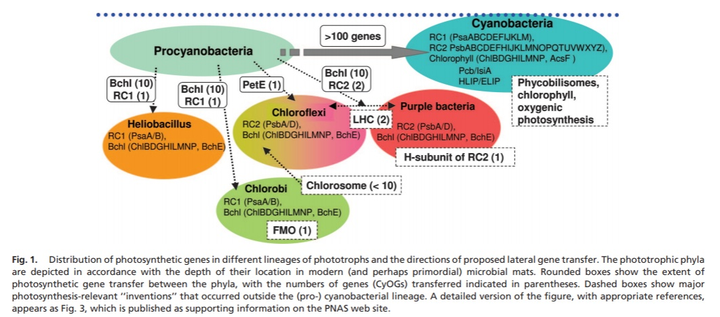
早期的光合细菌,便使用更易释放电子的有机物或硫化物作为电子来源。除蓝藻外,光合细菌主要包括紫色细菌和绿色细菌;与蓝藻和真核细胞不同,这些早期的不产氧光合细菌只有两个反应中心的其中一种。反应中心可能是从简单的结合叶绿素的膜蛋白进化而来,而这些蛋白原是为保护 DNA 免受紫外线的伤害而产生。在变形菌或绿弯菌门 Chloroflexi 中,I 型反应中心得以诞生,而 II 型反应中心则出现在日光杆菌 heliobacteria 或绿菌门 Chlorobi 中。
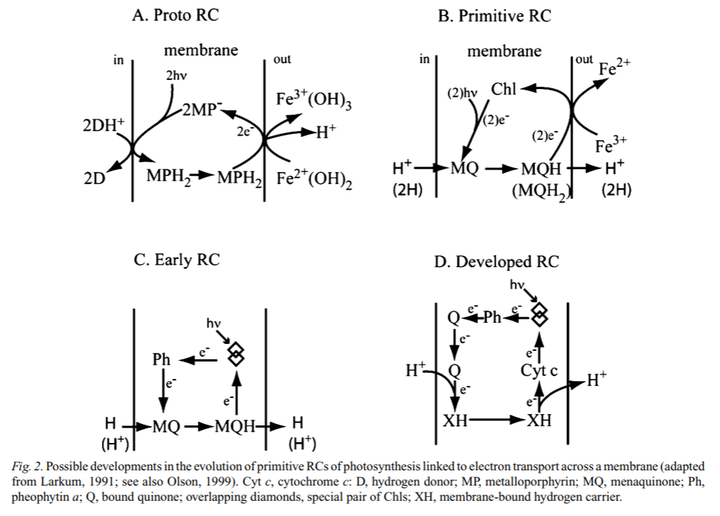
单独一种反应中心的能力有限,无法将电子从水中提取;而且,光合细菌没有叶绿素 a,只有效果较差的细菌叶绿素 a。这种物质来自于保护细菌免受紫外线辐射的色素,能够吸收光能,但想分解水还是超乎想象。因而,光合细菌只能利用有机物或还原的硫化物等作为电子源,将 CO2 还原成有机物,而不能利用环境中十分丰富的水资源。例如,紫色硫细菌和绿色硫细菌利用硫化氢为氢供体,在光下还原二氧化碳,而没有氧气的释放。
在非常早期的地球上,氢、氢气,硫化氢和甲烷等更容易释放电子的物质还容易获得,大量无氧的光合有机体也在那时开始发展。但在之后,可能是在 35 亿年左右,无氧光合微生物的生命活动大大降低了这些物质的含量。随着电子来源的耗尽,这些生物被迫适应更难提取电子的物质。这时,它们开始使用新的电子源:甲酸盐、碳酸氢盐和过氧化氢。
其中,过氧化氢可能是最主要的过渡电子源,它的分解难度在现有无氧光合细菌反应中心的能力范围内;但和氧气一样,过氧化氢对于无氧呼吸的细胞是一种致命的毒剂。它们会与还原性物质反应,产生羟基自由基来杀死细胞。古元古代雪球地球事件期间,强烈的阳光催生了大量的过氧化氢,由于过氧化氢的冰点略低于水(1°C),可以长期保存在冰中,当冰盖消散后,沉积的过氧化氢便释放到海洋和大气中,威胁附近的生物,使生物演化出各种过氧化物酶,以保护自身免受损伤。
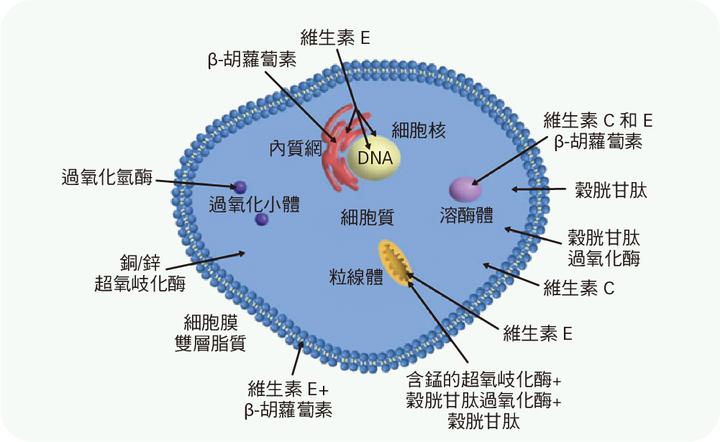
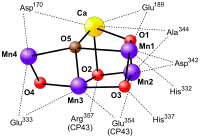
在此过程中,锰基过氧化氢酶从演化中产生。它具有双核锰金属中心,其结构类似于构成光合作用放氧复合物的四核锰金属中心,而这个四核锰金属中心可以将水氧化。因此,锰基过氧化氢酶可能就是这个水氧化中心的原型。在演化过程中,锰基过氧化氢酶开始被用于从过氧化氢中提取电子,释放出氧气,并将过氧化氢作为过渡性的电子源,继续着生命的进程;终于,二锰中心演化为四锰中心的水氧化中心,大大提升了氧化作用。
来自不同光合细菌的 I 型和 II 型反应中心也在随后融合,产生双反应中心系统。两种类型的反应中心的到来使得在两者之间建立了线性电子转移,从而可以从水中提取电子;同时,早期演化出的抗过氧化氢的酶也早在产氧光合作用进化之前进化,从而保护了第一个分解水产生氧气的光合生命。
当生物有了从水中提取电子而不被氧气损伤的能力时,有氧光合作用也正式产生。
有氧光合作用的出现要比无氧光合作用晚的多。大约 28-30 亿年前,这一历史性的突破发生在蓝藻的祖先中。26-27 亿年前,最早的确切蓝藻化石出现。它们从祖先的光合细菌中继承了它们的光合器官,并将它们保留下来。从此,生物开始大规模地利用阳光、水和二氧化碳作为原料来制造有机物质,将地球氧化,极大地促进了地球上的物质循环和能量转化。在随后的 23 亿年前,地球的氧含量开始上升;高效的有氧呼吸作用,为第一个线粒体内共生真核生物的进化点明了方向。
更久远的年代之后,某个真核细胞成功接纳了蓝藻,并由内共生产生了真核藻类。约 15 亿年前,植物的时代从此开始。从此,繁花似锦,郁郁葱葱。
当然,一切只是可能。