


现存的都不够像,有一类灭绝不算太久的鸟比较像,那就是恐鹤(Phorusrhacidae,也称为骇鸟 terror bird)。
但是说这一支完全灭绝了也不严谨,今天它的近亲也还生存着。
现生与恐鹤(Phorusrhacidae)关系最近的鸟类是叫鹤科(Cariamidae)。
目前仅存两个物种:红脚叫鹤(Cariama cristata)与黑脚叫鹤(Chunga burmeisteri),分布于南美洲中南部的草原与稀树草原。

在传统分类中,叫鹤曾被认为与鹤类近亲而分入鹤形目(Gruiformes),但最新的全基因组测序显示,它们实际上与已经灭绝的恐鹤科(Phorusrhacidae)关系更近,因此被独立归入叫鹤目(Cariamiformes)。
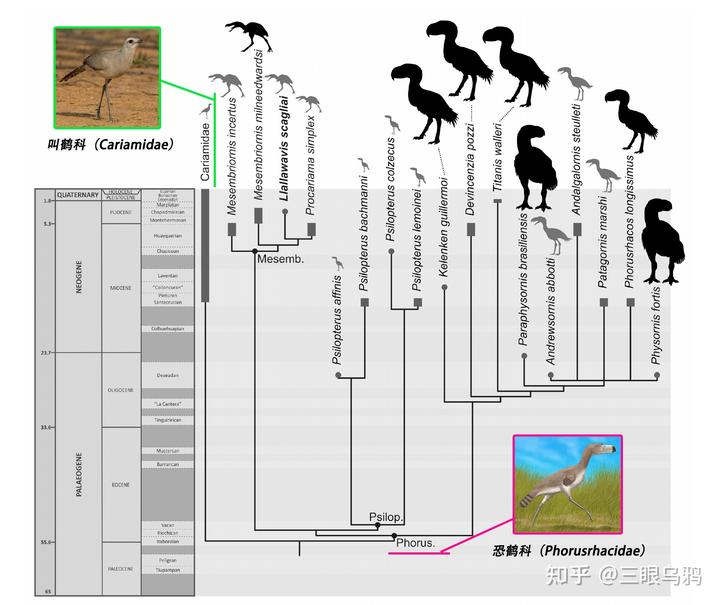
叫鹤不仅在系统发育上与恐鹤接近,在生态习性上也展现了许多相似点:
它们同样以地面活动为主,飞行能力有限。
双足奔跑迅速,依靠强壮的长腿和敏捷的机动性在开阔草原捕猎。
喙部锋利类似鹰隼,能够捕食昆虫、啮齿动物和小型爬行动物,遇到蛇类时甚至会用坚硬的喙和有力的双足将其击杀。
这些特征正是恐鹤的缩小版,算是鸟中豪杰的余脉。
至于会不会第三次复兴,那就要看天命了。

历史可以重演,但很难重复,覆灭的王朝终究不能再次掌权。
恐鹤(Phorusrhacidae)就是这样一个惊人的例子,它们几乎完成了恐龙未竟的事业,从肉食性、地面活动以及逐渐走向大型化这三个方面,都与中生代的兽脚类恐龙演化过程高度相似。
就像恐龙谱系中,从小型驰龙类一步步孕育出异特龙和霸王龙那样,恐鹤也在新生代经历了由小巧敏捷的奔跑捕食者到高达三米的巨型猛禽的演化历程。
它们的外形和兽脚类肉食恐龙更有惊人的相似之处:全部为双足奔跑,下肢骨骼粗壮结实,前肢退化为短小的翅膀,头颅却巨大,喙部弯曲如镰刀,攻击方式犹如巨斧劈下,强健的咬合力与迅猛的爆发力让人直接联想到霸王龙。
恐鹤的存在本身,就是恐龙遗族的再次巨型化(gigantism)*,几乎可以被看作恐龙王朝的二次复兴。
* 除了恐鹤之外,鸟类中还有多个巨型化的典型案例:恐鸟(Dinornithiformes)在新西兰演化为身高超过三米、体重两百公斤以上的植食性巨鸟;象鸟(Aepyornithiformes)在马达加斯加发展为近半吨重的庞然大物;古新世–始新世的巨型企鹅(如 Kumimanu、Palaeeudyptes)体长可达两米,远超现生企鹅。它们的祖先普遍是小型、善飞的鸟类,但在缺乏哺乳类竞争、生态位开放的环境中,逐渐演化为不会飞的巨型形态。
它们的故事始于约六千二百万年前的古新世,那是恐龙灭绝后的余波时期,哺乳类尚未在全球形成压倒性的掠食者体系,而孤立的南美洲为鸟类提供了前所未有的舞台。

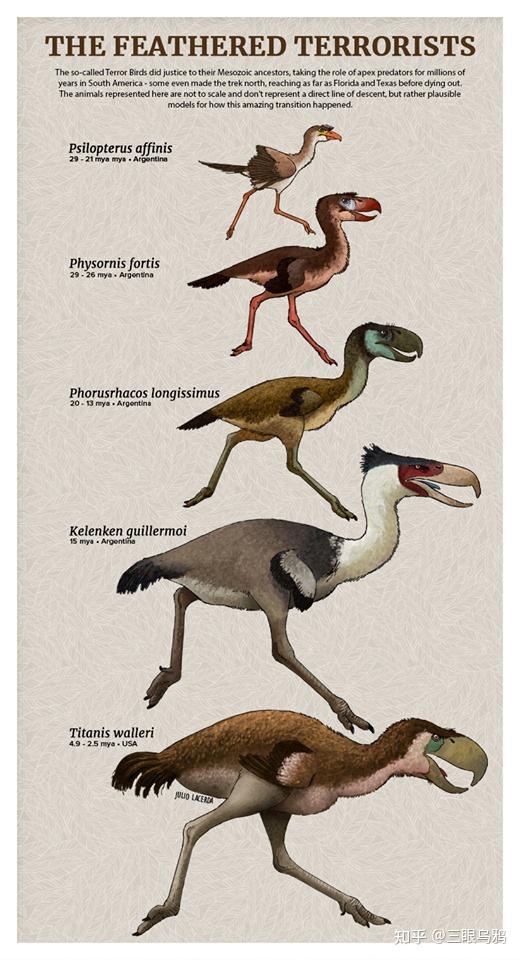
早期的代表如裸翼鸟(Psilopterus),体型不足一米,体重不到十公斤,行动敏捷,捕食小型哺乳动物和爬行动物。它们是典型的奔跑型猎手,也是恐鹤家族最古老且延续时间最长的分支。
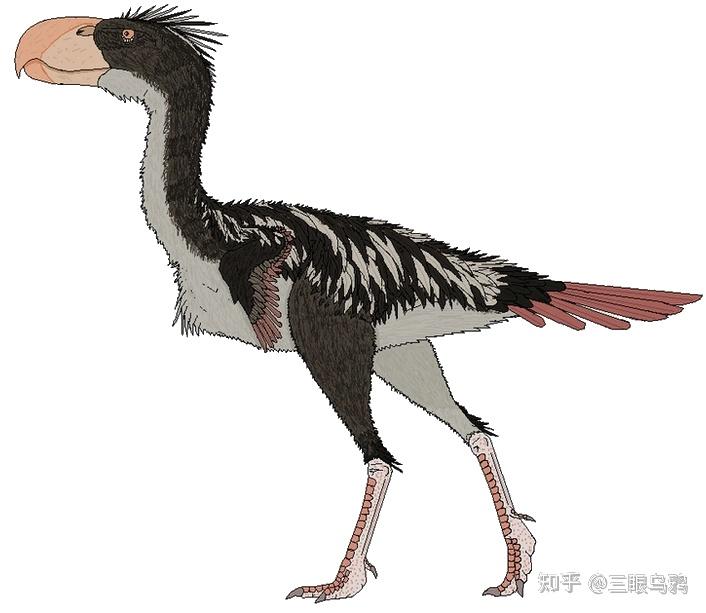
随着时间推移,这些早期小型猎手逐渐分化,中型种类如安达尔加鸟(Andalgalornis)和巴塔哥尼亚鸟(Patagornis)出现在渐新世和中新世,身高可达一米半到两米。
研究显示,它们的咬合力并不算极端强大,却依靠敏捷的上下劈击式攻击制造致命伤害,战术更像是一位持斧的屠夫。
在中新世中期,恐鹤迎来了真正的巨型化,这是它们最接近恐龙的时刻。
阿根廷发现的吉氏卡林肯窃鹤(Kelenken guillermoi)便是其中的巅峰,生活在约一千五百万年前,身高超过两米半,体重可达两百公斤。
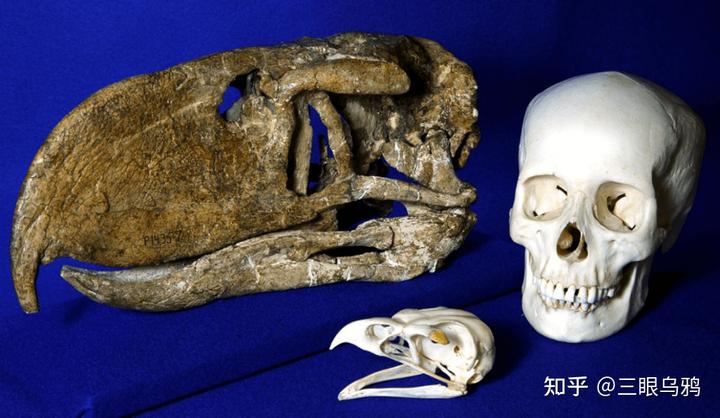
头骨长达七十一厘米,是已知鸟类中最长的头骨。

它在体型和生态位最接近犹他盗龙 (Utahraptor ostrommaysorum),都是各自环境中的顶级奔跑捕食者。

不过犹他盗龙毕竟是真正的兽脚恐龙,咬合力强健,牙齿尖锐,有着完整的尾巴和强壮前肢,爪子粗壮锋利。
它是已知体型最大的驰龙类之一,体重约 400-600kg(有保守估算约 280-300 kg,但多数学者认为较大标本体重可达 500 kg)。

拥有尖锐带锯齿的牙齿,非常适合撕裂猎物。
第二脚趾上的镰刀爪长约 22-24 厘米,是它捕猎的主要杀伤武器。全身骨骼粗壮且尾巴结构强劲,有助于在捕猎过程中稳定身体并释放爆发力。

虽然目前没有一个经严格肌肉重建确认的咬合力数值,但依据牙齿和颚部比例,以及与其同类恐龙的对比,可以推测其咬合力显著强于中小型兽脚类,足以对中大型猎物造成严重损伤。
与之相比吉氏卡林肯窃鹤只能算残血版恐龙。

另一巨型种类波氏德文森鸟(Devincenzia pozzi)出现在乌拉圭,体型同样庞大。
这些巨鸟在南美草原与林缘地带称霸,任何中型哺乳动物都可能成为它们的猎物。那一刻,恐鹤几乎重现了恐龙的辉煌:它们是新生代的大地霸王,重走了一条与兽脚类恐龙惊人相似的道路。

然而,在南美伊甸园顺利发育的恐龙遗族,温室终究要被打破。
大约三百万年前,巴拿马地峡隆起,南美洲与北美洲连通,掀起了美洲生物大迁徙。这是一个复杂的时代,南北美洲的物种开始双向交流,许多南美动物在新竞争格局下迅速衰落,但恐鹤家族的最后代表——泰坦鸟(Titanis walleri),却并非失败者。

它不仅没有坐以待毙,反而是迁徙中的成功北伐者之一,进入了北美,并在德克萨斯和佛罗里达留下了化石。这说明它在与北美掠食者的竞争中,至少在一段时间内站稳了脚跟,是恐鹤家族的最后一次辉煌。

它们的对手是刃齿虎属成员(如纤细刃齿虎Smilodon gracilis)、早期的美洲虎(Panthera onca)以及犬科掠食者的祖先。在这样的环境下,泰坦鹤依旧能保持一席之地,可见其适应力与竞争力之强。

尽管如此,恐鹤类最终还是没能熬过更新世的剧烈动荡。气候波动成为压垮它们的关键因素。当时地球进入冰期循环,北美的环境从开阔的稀树草原逐渐变为森林和湿地,泰坦鸟赖以捕猎的大型草食动物数量锐减或迁徙,它们的食物来源日益紧张。
生态位的萎缩并非孤立发生。就在更新世早期,哺乳类捕食者家族也在同步经历巨型化浪潮:剑齿虎家族中出现了体型更大的种类,如致命刃齿虎(Smilodon fatalis),逐渐占据了开放草原的伏击位点;犬科祖先扩张为强壮的群居奔跑猎手,为后来的恐狼奠定了基础;熊科中则出现了体型庞大的短面熊系成员,凭借力量和广阔的取食范围构成新的威胁。

这些新兴巨兽在不同生态层面切割了恐鹤赖以生存的空间,从草原上的高速追逐到林缘的埋伏伏击,几乎处处有哺乳类对手。泰坦鸟虽然凭借速度和巨喙一度立于不败之地,但随着环境转为森林与湿地,它们赖以捕食的大型草食动物逐渐减少,而竞争者却在不断增多。
最终,气候转型与哺乳类的全面扩张叠加在一起,令这一南美旧王朝的最后血脉在约一百八十万年前彻底走向终结。

回顾恐鹤的六千万年,它们几乎上演了恐龙的复兴,凭借巨型化再一次占据南美顶级掠食者的宝座。
它们从小型奔跑者进化为大陆霸主,走过了一条与兽脚恐龙高度平行的道路。
然而,恐龙王朝的二次登顶最终还是梦碎:泰坦鸟的北伐是家族的最后辉煌,却未能抵抗气候变化和新掠食者群体的合力冲击。
就像拿破仑二世的波拿巴王朝终究无法复辟,白垩纪大灭绝的王者遗族再也回不到舞台中央。
最终,这段辉煌像其他恐龙化石一样被埋入厚厚的泥土之下。
引用参考
Ameghino, H. F., & Alvarenga, H. M. F. (2003). Systematic revision of the Phorusrhacidae (Aves: Ralliformes). Papéis Avulsos de Zoologia (São Paulo), 43(4), 55–91. https://doi.org/10.1590/S0031-10492003000400001
Degrange, F. J. (2021). A revision of skull morphology in Phorusrhacidae (Aves, Cariamiformes). Journal of Vertebrate Paleontology, 40(6), e1848855. https://doi.org/10.1080/02724634.2020.1848855
Tambussi, C. P., de Mendoza, R., Degrange, F. J., & Picasso, M. B. (2012). Flexibility along the neck of the Neogene terror bird Andalgalornis steulleti (Aves: Phorusrhacidae). PLoS ONE, 7(5), e37701. https://doi.org/10.1371/journal.pone.0037701
MacFadden, B. J., Labs-Hochstein, J., Hulbert, R. C., Jr., & Baskin, J. A. (2007). Revised age of the late Neogene terror bird (Titanis) in North America during the Great American Interchange. Geology, 35(2), 123–126. https://doi.org/10.1130/G23186A.1
Blanco, R. E., & Jones, W. W. (2005). Terror birds on the run: A mechanical model to estimate its maximum running speed. Proceedings of the Royal Society B: Biological Sciences, 272(1579), 1769–1773. https://doi.org/10.1098/rspb.2005.3133
LaBarge, T. W., Gardner, J. D., & Organ, C. L. (2024). The evolution and ecology of gigantism in terror birds (Aves, Phorusrhacidae). Proceedings of the Royal Society B: Biological Sciences, 291(20240235), 1–9. https://doi.org/10.1098/rspb.2024.0235
Woodburne, M. O. (2010). The Great American Biotic Interchange: Dispersals, tectonics, climate, sea level and holding pens. Journal of Mammalian Evolution, 17(4), 245–264. https://doi.org/10.1007/s10914-010-9144-8